№7, 2006 г.
© Хлебович В.В.Новое окно в эпигенетику
В.В. Хлебович
Владислав Вильгельмович Хлебович, доктор биологических наук, профессор, главный научный сотрудник Зоологического института РАНЭйфория, вызванная полным раскрытием генома многоклеточных (и даже человека!), сменилась недоуменной паузой. А что дальше? Стало ясно, что от знания состава генома до управления фенотипом дистанция очень велика (как от набора книг военных уставов до создания реальной армии). В этой связи все чаще стало звучать слово "эпигенетика" (от греч. epi - над, после) - так в 1947 г. К.Х.Уоддингтон предложил называть "ветвь биологии, изучающую причинные взаимодействия между генами и их продуктами, образующими фенотип". Сейчас эпигенетика - широкое понятие, отражающее онтогенетические, физиологические, молекулярные и эволюционные аспекты регуляции активности генов [1, 2].
Однако лаконичность и образность научных терминов таят в себе опасность их заимствований другими науками, искажению и размыванию начальных понятий. Не так давно происходило это с экологией: очень соблазнительное, лежащее в основе термина слово "экос" (от греч. oicoV - дом), породило яркие, но явно ненаучные словосочетания - экология культуры, экология производства, экология половой жизни и т.д. Похоже, нечто подобное грозит эпигенетике, на что жаловался еще сам Уоддингтон в 1970 г., настаивая на сохранении изначальных рамок понятия.
В последнее десятилетие развитие эпигенетики связано с большими успехами в исследованиях молекулярных механизмов процессов экспрессии-репрессии генов - путем метилирования-деметилирования цитозина и последующей активной роли хроматина [2]. Появилось представление об эпигенетической наследственности - передаче через митозы модифицированного цитозином состояния ДНК. Возникло даже новое определение эпигенетики: "исследование митотических и/или митотически наследуемых изменений в функционировании генома, которые не могут быть объяснены изменениями в последовательности ДНК" [3]. Положение, очевидно, справедливо, но все же сужение рамок эпигенетики только до особенностей молекулярных механизмов уводит в тень традиционный для отечественной биологии подход к изучению эволюции.
Дискретные адаптивные нормы
Со времен Ч.Дарвина изменчивость подразделяют на неопределенную, или наследственную, т.е. затрагивающую генотип, и определенную, при которой меняется только фенотип (преобразуется форма) практически всех особей одной популяции (породы, сорта), реагирующих на определенное внешнее воздействие.
Современные и наиболее привычные синонимы определенной изменчивости - морфозы, модификации и широко используемая с легкой руки Г.Ф.Гаузе [4] в англоязычной литературе фенотипическая пластичность (phenotypic plasticity). Определенная изменчивость может оказаться инадаптивной, на практике часто вызываемой искусственными химическими или физическими воздействиями (их результат обычно называют морфозами) или явно адаптивными в ответ на изменение природного фактора среды или воздействия, имитирующего этот фактор. В последнем случае такая изменчивость называется адаптивной модификацией, фенотипическим окном генома или адаптивной нормой. Среди адаптивных норм (термин И.И.Шмальгаузена) Б.М.Медников выделил дискретные адаптивные нормы (ДАН) - резко, без переходов, различающиеся по принципу "да или нет", т.е. каждое изменение четко адаптировано к определенному воздействию [5].
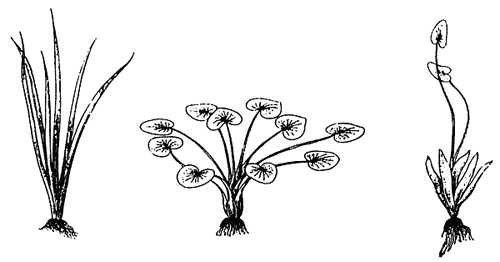
Три вида стрелолиста.
У североамериканского стрелолиста округлого (а) все листья подводные, в виде цилиндрических, поперечно-перегородчатых черешков без пластинок. Тропический афроазиатский стрелолист цепкоплодный (б) обладает только плавающими листьями с сердцевидными пластинами. У широко распространенного в России стрелолиста обыкновенного (в) на поверхности воды стреловидные листья, а под водой - линейные.Примеры ДАН всем хорошо известны из школьной программы. Самые яркие из них - морфы стрелолиста (Sagittasria) с разной (в зависимости от условий обитания) формой листьев, крылатые и бескрылые формы насекомых, возникающие в ответ на плотность популяций и др. Различия между морфами иногда бывают поразительными, особенно когда они отмечаются между особями одного клона, т.е. имеющими одинаковый геном. В клонах некоторых инфузорий, питающихся микроорганизмами, при недостатке корма образуются две морфы - гиганты и карлики. Карлики довольствуются малым количеством пищи, а гиганты питаются собратьями.

Гигантская (200-300 мкм) особь инфузории Onychodromus quadricornutus заглатывает карлика (показано стрелкой), собрата по клону [6].
Некоторые наездники-энциртиды откладывают яйца в тело насекомых, где в большом объеме питательной массы это яйцо подвергается полиэмбрионии - первичные бластомеры расходятся и каждый образует новую особь. Таким образом создается клон личинок. В этом клоне у некоторых немногочисленных особей необычно разрастаются жвалы и существенно меняется поведение. Такие личинки истребляют конкурентов-наездников не только чужого, но и своего вида, которые посмели покуситься на занятое клоном пространство. Специализация этих личинок-охранников зашла так далеко, что они утратили способность линять и превращаться во взрослое насекомое, способное давать потомство. Такие случаи морф-камикадзе, естественно, могли сформироваться только в результате группового отбора.
Очевидно, помимо ДАН существуют и градуальные адаптивные нормы (ГАН), которые реализуются постепенно - в зависимости от дозы модифицирующего фактора. Например, у растений снижение освещенности приводит к постепенному увеличению листовой пластинки, у некоторых животных в зависимости от обеспеченности пищей меняется масса тела и время полового созревания.
С большой долей уверенности можно считать проявления ДАН (а возможно, и ГАН) результатом регуляции активности генов, т.е. эпигенетическим процессом, или, по Медникову, экспрессионным полиморфизмом. А.С.Серебровский предложил еще более удачный, по нашему с Ю.А.Лабасом мнению [7], термин - фенотипическое окно генома [8].
ДАН и наследственные триады
Наше внимание привлекли виды (популяции, штаммы) организмов, которые не только способны образовывать ДАН, но и часто имеют близких "родственников" с наследственной формой, т.е. фиксированной морфой. Чтобы легче было понять, о чем речь, приведем несколько примеров.
Среди растений, помимо уже упомянутых видов стрелолистов с разными листьями, известны постоянно наземные или постоянно водные виды лютиков, а также очень близкие им виды с соответствующим переключением ДАН. У китайской примулы (Primula sinensis) в зависимости от температуры и влажности могут образовываться цветы с белой и красной окраской, а кроме того, существуют постоянные формы, никогда не меняющие окраску лепестков.
Триада рас китайских примул. Диморфизм свойствен и некоторым грибам - в зависимости от обеспеченности ресурсами они способны расти в форме мицелия или дрожжей, при этом есть штаммы тех же видов, которые независимо от условий существуют всегда в одной из этих форм.
В клоне некоторых коловраток из рода Asplanchna могут одновременно присутствовать и мелкие, и крупные формы, что позволяет им более эффективно использовать пищевые ресурсы. Кроме того, известны и очень близкие формы (виды), у которых развиваются только крупные или только мелкие особи.
Похожая на саламандру кротовидная амбистома (Ambystoma talpoideum), адаптируясь к размерам водоема, может размножаться или на стадии личинки, или на стадии, прошедшей метаморфоз. Помимо того, существуют особи с наследственно фиксированной стадией размножения.
Даже несколько этих примеров наводят на мысль, что виды, способные к образованию ДАН, часто организованы в наследственные триады с двумя формами, отвечающими фиксированным морфам диморфного таксона.
Неоднократное подтверждение этой закономерности породило гипотезу: в основе способности к образованию ДАН лежит существование минимум двух включаемых разными факторами среды альтернативных наследственных программ (A B). Два других члена триады (А и B) возникают в результате наследственной фиксация флангов этой широкой адаптивной нормы, причем их образование, очевидно, связано с выпадением (инактивацией) одной из альтернативных программ.
Оборонительные морфы
О присутствии в среде хищника жертва узнает по выделяемым им сигнальным веществам (кайромонам), что вызывает у организмов-жертв морфологические изменения (увеличиваются размеры и меняется форма тела, образуются особые лопасти или шипы и т.д.), не позволяющие их схватить и съесть. Такие морфы (типичные ДАН) называют соответственно их адаптивному значению оборонительными. Они весьма разнообразны. Одноклеточные водоросли при появлении хищных коловраток или дафний резко увеличиваются в размерах. У некоторых инфузорий в присутствии хищных червей расширяется тело, на котором разрастаются боковые лопасти. При появлении в толще воды хищных коловраток Asplanchna на панцирях их жертв, мирных коловраток Brachionus и Keratella, образуются мощные шипы. У нескольких видов дафний образуются затылочные шипы, если они почувствовали хищных личинок комаров Chaoborus или клопов Notonecta. У серебряных карасей расширяется тело при подсадке в их водоем щуки или окуня; образование такой оборонительной морфы у карасей, описанное всего десятилетие назад, удивило даже специалистов-ихтиологов.
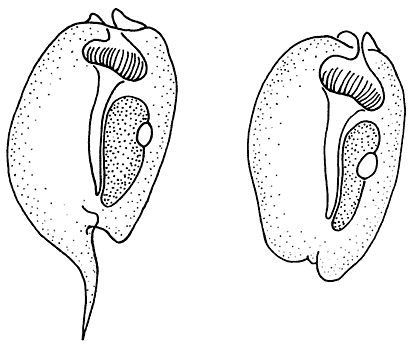
Оборонительная (слева) и типичная формы
инфузории Entodinium caudatum из желудка козы.
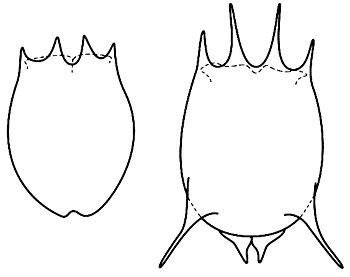
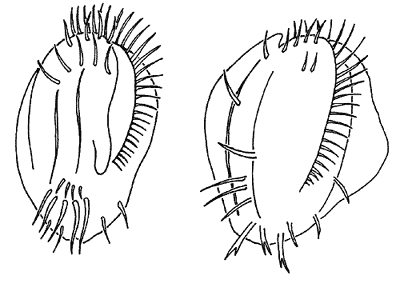
Здесь и далее прорисовки Ю.Р.ОхлопковаПанцири типичной (слева) и оборонительной форм
коловратки Brachionus calyciflorus.Очевидно, что оборонительные морфы - некий компромисс между выигрышем в выживании и гидродинамическим проигрышем (с выростами плавать труднее). Поэтому при исчезновении признаков присутствия хищника потенциальная жертва возвращается к типичному для нее облику.
Примечательно, что все без исключения виды, способные к образованию оборонительных ДАН, относятся к таксонам, размножающимся всегда или значительную часть времени (в самое продуктивное - летнее) бесполым путем (партеногенетически). К таковым относится и гиногенетический (от греч. gunh - женщина) серебряный карась, у которого нет собственных самцов, участие же в размножении самцов чужих видов весьма условно - их сперматозоиды лишь активируют развитие яйца, не объединяясь с его ядром. Все преобразования во всех упомянутых группах организмов происходят на уровне клонов, т.е. среди особей с идентичным геномом, а, следовательно, образование оборонительных ДАН - в чистом виде эпигенетический процесс. Это знание дает исследователям чрезвычайные методические преимущества: уверенность в генетической однородности материала и возможность многократного воспроизведения эксперимента. К тому же можно добавить относительную простоту культивирования клонов жертв, многие из которых широко используются как лабораторные культуры (одноклеточные водоросли, инфузории, коловратки, дафнии).
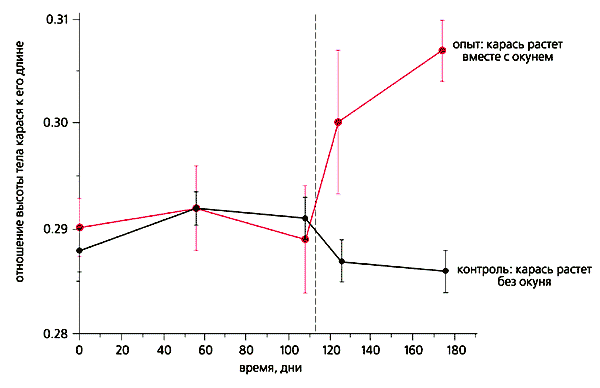
Образование оборонительной формы серебряного карася в ответ на хищничество окуня.
Вертикальным пунктиром отмечено время, когда окуня переключили с питания беспозвоночными на питание карасем [12].Для проверки гипотезы (которая основана, напомним, на предположении, что образование ДАН связано не с включением-выключением одного или блока генов, а существованием двух альтернативных наследственных программ) мы использовали клоны организмов, способных к образованию оборонительных морф, - простейшей зеленой водоросли Scenedesmus aсutus, одноклеточной брюхоресничной инфузории Euplotes aediculatus и многоклеточной дафнии Daphnia pulex. (Эта эксперименты выполнены вместе с аспирантом А.В.Дегтяревым.)
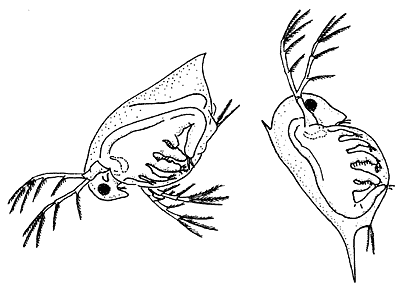
Типичная (слева) и оборонительная (реакция на витамин Е) формы дафнии Daphnia pulex.
Превращение одной формы в другую блокируется актиномицином D.В качестве ингибитора экспрессии генов выбран актиномицин D (AmD), широко используемый именно для этих целей в биологии развития уже около полувека. Как ингибитор ДНК-зависимой РНК-полимеразы он специфически подавляет транскрипцию иРНК de novo. Иными словами, если какой-либо процесс будет подавляться нетоксичными дозами AmD, это будет доказательством того, что запускается он благодаря экспрессии "своего" гена (группы генов).
К брюхоресничной инфузории подсаживали питающихся ими турбеллярий Stenostomum leucops. Непрерывно размножаясь, инфузории под влиянием хищника постепенно становились шире, но добавление нетоксичной дозы AmD подавляло этот процесс, и форма тела инфузорий в присутствии хищника оставалась как в контроле (без турбеллярий). Когда из среды со сформировавшейся оборонительной морфой инфузории убирали хищника, форма ее тела возвращалась к типичной, но в этом случае AmD подавлял процесс, и морфа фиксировалась как оборонительная даже без хищника!
Типичная и оборонительная (ответ на подсаживание хищной турбеллярии Stenostomum) морфы инфузории Euplotes aediculatus.
Превращение одной формы в другую полностью блокируется актиномицином D.Клетки зеленой водоросли в присутствии хищников - коловратки Branchionus calyciflorus или дафнии Daphnia pulex - резко увеличивались в размерах, что можно было остановить добавлением AmD. После удаления из среды беспозвоночных ставшие под их влиянием гигантскими клетки водоросли постепенно, деление за делением, возвращались к исходному размеру, но и этот процесс блокировался AmD.
Образование оборонительных морф у дафнии изучали многие специалисты, но обычно на нее "натравливали" личинок комара Chaoborus, в результате чего у дафнии позади головы появлялся оборонительный шип. Из-за отсутствия личинок комара мы решили добавить в среду a-токоферол (витамин E) - метаболит хищной коловратки Asplanchna, служащий сигналом для ее жертв - коловраток Brachionus и Keratella - превращаться в оборонительную шипастую форму. Однако никто не проверял, как подействует a-токоферол на дафний. Результат оказался неожиданным: шипы появились, но на противоположном конце тела. Дафнии с каудальным (хвостовым) шипом довольно обычны в наших прудах и даже отмечалось, что эти формы более доступны для личинок комара, чем обычные (без шипов). Вероятно, в природе молодь столь мелкой дафнии, как Daphnia pulex, может быть жертвой хищных коловраток, отсюда и реакция их на a-токоферол. Кроме того, напрашивается и более важный вывод: у одного и того же вида (в данном случае Daphnia pulex) могут быть разные проявления оборонительных морф в зависимости от особенностей хищника (в данном случае личинки комара вызывали образование затылочного шипа, а метаболит коловратки - каудального шипа).
Раз уж мы заговорили об обобщениях, то возникает естественный вопрос: можно ли проводить аналогию между процессами образования оборонительных морф у простейшей водоросли, одноклеточного (инфузории) и многоклеточного (дафнии) животных? Безусловно, дело лишь во времени. Одноклеточные, размножаясь ежедневно, начинают модифицироваться в нужном направлении практически сразу после внешнего воздействия (сигнала хищника). А у многоклеточной дафнии, у которой от яйца до яйца проходит около десяти дней, признаки адаптивного преобразования тела начинает проявлять спустя одну-две недели после добавления в среду a-токоферола. Как и у одноклеточных, у дафнии AmD блокирует возникновение оборонительных структур. Если же перестать добавлять a-токоферол, то через одну-две недели у потомков видоизмененной дафнии шипы начнут уменьшаться и в конце концов исчезнут вовсе. AmD блокирует и этот процесс.
Таким образом, во всех трех случаях актиномицин D подавлял как адаптивное образование оборонительной морфы, так и возвращение особей клона к типичному состоянию после удаления хищника или "сигнала тревоги". Это - свидетельство того, что за каждой морфой стоит реализация собственной наследственной программы. Конечно, эти обе программы у способных к образованию ДАН организмов функционально тесно связаны - прежде, чем включится одна, должна выключиться другая. В этом отношении их тандем может считаться эпигеном, поскольку "эпигеном называют наследственную единицу (циклическую систему), имеющую не менее двух режимов работы подчиненных ей генов и способную сохранять каждый из режимов в последующем ряду поколений" [1]. Теперь есть все основания утверждать, что виды, способные к образованию дискретных адаптивных морф, имеют соответственно этому более богатый геном.
ДАН и эволюция
Роль модификационной изменчивости в эволюции обсуждается более века. Н.П.Дубинин и В.Грант категорически утверждали, что поскольку по определению модификация происходит при неизменном геноме, ее адаптивное проявление уводит организмы от действия отбора и фактически тормозит эволюционный процесс. Логично, не правда ли?
Другая точка зрения разделялась большинством не менее авторитетных эволюционистов ушедшего века (Х.Ф.Осборн, Дж.М.Болдуин, В.С.Кирпичников, Е.И.Лукин, И.И.Шмальгаузен, Г.Ф.Гаузе, К.Х.Уоддингтон, Ю.И.Полянский). Видимо, все они считали свои взгляды оригинальными, особенно в части описаний форм естественного отбора, при этом были предельно корректными и доброжелательными друг к другу, что для наших соотечественников было особенно важно и ценно в тяжелые 30-50-е. Эти позиции, казавшиеся тогда различными, сейчас выглядят как общая, которую четко сформулировал в 1984 г. Гаузе: "Адаптивная модификация, будучи продуктом естественного отбора, уже в силу своей адаптивности указывает на то, что случайно возникшая похожая на нее мутация будет подхвачена отбором и будет эволюционно закреплена". Происходит закрепление в результате отбора, получившего в разное время разные названия - совпадающий отбор, косвенный отбор, генетическая ассимиляция, стабилизирующий отбор, изменение порога реагирования.
Думаю, к этой позиции близки и сторонники "эпигенетической теории эволюции" (М.А.Шишкин, Д.Л.Гродницкий, А.П.Расницын), считающие, что эволюционные преобразования начинаются с возникающих в ответ на изменения в среде морфозов, которые в случае их адаптивности подвергаются генетической ассимиляции и становятся наследственно закрепленными. Естественно, подразумевается, что модификационный признак и генетически ассимилированный сходны, но не тождественны, и за ними стоят разные группы генов.
В основе третьей точки лежит доказанное Жакобом и Моно существование аппарата регуляции активности генов у бактерий. Признание этого явления у эукариот, независимо от механизмов его осуществляющих, должно привести к ревизии важнейших эволюционных постулатов [7]. Гены могут быть в активном и неактивном (спящем) состоянии. Модификационная изменчивость связана с регуляцией активности генов. Отбор совершается только по признакам, проявленным в фенотипе, т.е. по активным генам. Регуляция активности генов открывает воздействию отбора только те гены, которые нужны для закрепления данного признака (адаптации). Если гены будут в спящем состоянии в течении многих поколений, в них могут накапливаться неподверженные отбору мутации. Эти мутации будут резервом нужной для эволюции неопределенной изменчивости в случае открытия соответствующего фенотипического окна генома при возвращении прежних условий. Очевидно, что открытие фенотипического окна генома через многие поколения его пребывания в спящем состоянии, учитывая периодические пульсации климата и реверсию границ экосистем, факт нередкий. Отсюда проистекает наше представление о направленном отборе - кажется, что среда сама открывает нужное направление изменчивости и эволюции.
Фенотипическое окно генома, очевидно, включает в себя несколько генов. Можно представить себе переход гена в другую генорегуляторную группу другого фенотипического окна в результате ассоциирующего отбора. Такую возможность в свое время мы проверили экспериментально [9]. Как известно, дрожжи Saccharomyces cerevisiae имеют два фенотипических окна генома - аэробное и анаэробное. В качестве исходного был взят штамм CrpR, устойчивый к аминазину (хлорпромазину) - ингибитору флавиновых ферментов, причем это свойство проявлялось как в аэробной, так и в анаэробной фазе. После длительного культивирования этих дрожжей в аэробных и анаэробных условиях мы получили своего рода триаду наследственных форм - исходную, устойчивую к аминазину во всех случаях, и еще две, каждая из которых проявляла это свойство только в определенных условиях. Это ли не свидетельство относительной легкости преобразования генорегуляторных комплексов.
Эпигенетический вариант эволюции
Начало формирования модификационной изменчивости, которая возникает в результате создания двух альтернативных наследственных программ, скорее всего связано с дупликацией генов и с последующим расхождением признака. Расхождение не должно быть симметричным. Один дупликант, видимо, должен быть неизменным, консервативным, чтобы гарантировать сохранение старых, проверенных в поколениях связей со средой - на всякий случай. Второй - "под прикрытием удвойки" (выражение А.С.Серебровского) может стать объектом эксперимента природы по освоению новой среды. Только так, например, можно представить формирование морф стрелолиста, независимо от того, какую из них следует признать первичной - водную или воздушную.
Дуплицироваться могут не только отдельные гены, но и целиком геном, в результате чего может возникнуть организм с четырьмя наборами хромосом (тетраплоид), что позволяет ему приспособиться к качественно новым условиям среды. Подтверждение тому было найдено при сравнительном анализе геномов двух близкородственных видов дрожжей - тетраплоида S.cerevisiae (с 16 парами хромосом) и диплоида Kluyveromyces waltii (с 8 парами хромосом) [13, 14]. Выяснилось, что многие их гены гомологичны, при этом каждой паре хромосом K.waltii соответствует две пары S.сerevisiae. Из этого следует, что произошли они от общего диплоидного предка. Удвоение его генома привело к возникновению S.сerevisiae, у которого за 100 млн лет каким-либо изменениям подвергся лишь один ген из каждой пары, второй же оставался неизменным, очевидно, "подстраховывая" возможные преобразования первого.
Так называемую наследственную фиксацию признака, который ранее проявлялся как модификационный, легче всего объяснить выпадением (дезактивацией) альтернативной программы. Этот акт стабилизации признака, ранее признававшийся Шмальгаузеном и Гаузе важнейшим прогрессивным моментом эволюции, на самом деле, очевидно, связан с потерями в геноме, скорее всего с делециями. При этом наследственно фиксируется не просто сходный по своим проявлениям с модификацией признак, как считали классики, а признак ему абсолютно идентичный.
В природе эпигенетическая изменчивость в чистом виде, наиболее доступном исследованию, проявляется в трех случаях:
- в каскаде клеточных дифференцировок (потомки одной зиготы);В основе эпигенетических процессов лежат альтернативно (ДАН) или последовательно (онтогенез) активированные наследственные программы (гены, блоки генов). Способные к таким процессам организмы имеют соответственно более богатую наследственную программу. Обогащение организмов наследственными программами, очевидно, составляет часть эволюционного прогресса.- в образовании дискретных адаптивных норм у клональных организмов;
- в повторных акклимациях к природным факторам, когда один и тот же организм, естественно, при одном и том же геноме, может многократно повторять сценарии экспрессии-репрессии генов.
Адаптивные модификации не что-то, неизвестно откуда взявшееся и потом "генетически ассимилированное", а результат предварительного удвоения и дивергенции конкретной наследственной программы.
Есть основания полагать, что через эпигенетическую эволюцию прошли некоторые таксоны рангом выше вида и рода.
В результате такой эволюции образовались триады форм, одна из которых с широкой нормой реакции (ДАН), а две других представлены ее наследственно фиксированными флангами. Этот процесс может повторяться многократно.
Например, у многих клопов-водомерок семейства Gerridae существуют крылатые и бескрылые ДАН как адаптивная реакция на размеры водоема: из маленькой грозящей пересыханием лужи нужно улететь (сигнал - стресс от столкновений), а в большом озере нет смысла тратиться на образование крыльев и полет. Кроме того, среди водомерок много видов, которые могут быть либо только крылатыми, либо только бескрылыми. Эволюция в семействе клопов-водомерок шла от видов с широкой нормой реакции (с ДАН) к видам с фиксированными морфологическим приобретением (с крыльями), от которых впоследствии образовались новые виды с модификационной изменчивостью [10].
Другой пример - эволюция видов большого подсемейства морских окуней (Sebastinae) [11]. На больших пространствах двух океанов, Атлантического и Тихого, было семь синхронных циклов видообразования, которые оказались связанными не с классической географической изоляцией, а с биотопической. В каждом цикле один старый вид, обычно из средних глубин, сохраняя себя, порождал два новых вида - один мелководный, другой глубоководный. Так образовывались тройки родственных видов. Затем спустя некоторое время все повторялось в следующем цикле.
Очевидно, картины эволюции таких удаленных друг от друга групп, как клопы-водомерки и морские окуни, подтверждают наши представления об эволюционной роли ДАН.
Литература
1. Голубовский М.Д., Чураев Р.Н. Динамическая наследственность и эпигены // Природа. 1997. №.4. С.16-25.
2. Карпов В.Л. От чего зависит судьба гена // Природа. 2005. № 3. С.34-43.
3. Bird A. // Genes. Dev. 2002. V.16. №1. P.6-21.
4. Гаузе Г.Ф. Экология и некоторые проблемы происхождения видов // Экология и эволюционная теория. Л., 1984. С.5-105.
5. Медников Б.М. // Журн. общ. биол. 1987. Т.43. №1. С.15-26.
6. Wicklow B.J. // J. Protozool. 1988. V.35. №1. P.137-141.
7. Лабас Ю.А., Хлебович В.В. "Фенотипическое окно" генома и прогрессивная эволюция // Соленостные адаптации водных организмов. Л., 1976. С.4-25.
8. Серебровский А.С. Некоторые проблемы органической эволюции. М., 1973.
9. Михайлова Н.П., Лабас Ю.А., Сойдла Т.Р., Хлебович В.В. Мутации устойчивости к аминазину, специфически проявляющиеся при разных типах энергетического обмена дрожжевых клеток // Соленостные адаптации водных организмов. Л., 1976. С.232-236.
10. Andersen N.M. // Oikos. 1993. V.67. №3. P.433-443.
11. Барсуков В.В. // Вопросы ихтиологии.1981. Т.21. №1. С.3-27.
12. Brцnmark Ch., Petterson L.B. // Oikos.1994. V.70. P.396-402.
13. Wolfe K.H., Schields D.C. // Nature. 1997. V.387. P.708-713.
14. Kellis M. et. al. // Nature. 2004. V.428. P.617-624.