Дарвиновская теория эволюции исходила во многом из аналогии между процессами, происходящими в природе, и селекцией домашних животных и растений. Селекционер был заменен условиями обитания - от них зависят скорости успешного размножения (эта краткая формулировка, конечно, неполна, так как живые существа меняют среду обитания и сами составляют ее важный элемент). Не так просто заметить связанные с этим явления в природе - они обычно протекают медленно. Но человек все чаще вторгается в окружающую среду. В худшем случае это приводит к исчезновению многих ее обитателей, а иногда - к их заметному изменению. Говорят, что живые существа, видоизменяясь, приспосабливаются к новым условиям.
Термин ``приспособление'' метафоричен и неточен. Он был бы более уместен в том случае, когда изменения происходили бы целенаправленно и сознательно. В действительности же наиболее важен такой механизм: одним обитателям становится существенно хуже, другим - не настолько (или даже лучше), и эти последние вытесняют прежних жителей, даже если вначале, до изменения в среде, были в меньшинстве. Называя этот процесс отбора приспособлением, мы как бы смешиваем его с другими - индивидуальной адаптацией и модификацией индивидуального развития. При изменении условий все организмы так или иначе меняются - адаптируются. Это происходит довольно быстро, но в сравнительно узких пределах. Более широкий диапазон изменений дает модификация индивидуального развития. В новых условиях развитие идет хоть немого, да по-другому. Для проявления таких модификаций требуется уже смена нескольких поколений, хотя бы одного-двух. Эти изменения обычно обратимы и захватывают сразу значительную часть особей. Не вытеснение одних другими за счет изменений в коэффициенте размножения, а просто изменение многих - в этом отличие от эволюции путем отбора наследуемых вариаций. Он действует медленнее всех индивидуальных приспособлений и модификаций, но зато может приводить к ``приспособлениям'', намного сильнее уклоняющимся от первоначальных свойств.
Указанные ``приспособления'' поставляют много материала для изучения эволюционных изменений. Один из примеров был описан английским зоологом В. Уэлдоном. Наше изложение следует вступительной статье К. А. Тимирязева к книге Ч. Дарвина [38].
В. Уэлдон изучал мелкого краба, водящегося в Плимутской бухте. Там был построен большой мол. Перегораживающий вход в бухту. После этого в фауне бухты обнаружились изменения. В. Уэлдон измерял в числе прочих показателей ширину лобной части головогруди краба (человек, видевший краба, поймет почему говорится ``головогрудь'', а не отдельно голова и грудь). Он заметил, что ширина эта год от года уменьшается. Связав это с постройкой мола, он предположил, что все дело в мутной воде, и провел опыты в аквариумах с водой разной чистоты. Как и следовало ожидать, в мутной воде смертность крабов была выше, причем широколобые крабы были более чувствительны к чистоте воды.
Итак, постройка мола вызвала замутнение воды, замутнение повысило смертность крабов, но по-разному: для широколобых сильно, для узколобых - послабее. В. Уэлдон пошел дальше и нашел физиологическую причину таких различий - у широколобых крабов хуже обеспечена фильтрация воды от ила и жабры загрязняются сильнее.
На резонный вопрос, почему же, если узколобые крабы такие ``хорошие'', они не вытеснили широколобых раньше, можно дать такой ответ: в нормальных условиях широколобые имели какие-то свои преимущества, а катастрофическое для них положение, создавшееся после постройки мола, свело эти преимущества на нет.
А вот следующий вопрос не так прост: откуда взялись узколобые крабы? Если они были до постройки мола, то почему их не вытеснили окончательно имевшие тогда преимущество широколобые? А если их не было, то как они возникли? Заданный вопрос об изменчивости - один из центральных в теории эволюции. Мы займемся им отдельно, а пока лишь отметим, что можно объяснить возникновение узколобых крабов без привлечения гипотезы о специальном механизме целесообразного реагирования на изменение среды.
Одна из самых знаменитых бабочек - скромная березовая пяденица. Она встречается в учебниках и монографиях чаще, чем красавец махаон. Обычная ее окраска - белая с серыми пятнышками. Днем она сидит, раскинув крылья, на стволе дерева, а активна ночью. Днем сидящих бабочек охотно поедают птицы, если, конечно, найдут - ствола обычно покрыты лишайниками тоже белыми с серыми пятнышками. Не так-то легко обнаружить на этом фоне березовую пяденицу.
Но промышленная революция привела к загрязнению воздуха и гибели лишайников. Стволы потемнели. И вот, около ста лет назад в Англии начали находить другую форму березовой пяденицы с серо-черными крыльями. С годами эта редкая вначале форма становилась все более многочисленной.
Эксперимент показал, что если в загрязненном лесу выпустить одинаковое число светло- и темно-окрашенных бабочек, то птицы будут съедать светлых в два-три раза чаще ([94], c, 47). Результат может показаться заранее очевидным, но как ни странно, дискуссия о роли защитной окраски у насекомых проходила в свое время настолько бурно, что эксперимент был необходим.
На примере спора о приспособительном значении окраски можно убедиться в том, что количественный язык необходим в биологии. Некоторые ученые отрицали пользу, приносимую защитной окраской ее обладателю, на том основании, что едят и тех и других, и хорошо, и плохо замаскированных. Но никто не утверждал, что защитная окраска гарантирует бессмертие. Важно количественное соотношение.
Проиллюстрируем это простой оценкой. Рассмотрим две разновидности бабочек,
одинаковые во всех отношениях, кроме окраски. Будем называть их бабочка А и
бабочка Б. Пусть из 100 бабочек А птицы за год съедают 50, из 100 бабочек Б
- 49, а вероятности гибели бабочек А и Б от других причин одинаковы. Если в
начальный момент числа бабочек А и Б равны, то через год их отношению (Б/А)
будет в среднем
![]() , через два года -
, через два года -
![]() , через
, через ![]() лет -
лет -
![]() .
.
В школьном курсе мельком встречается замечательное число
![]() .
Оно определяется как предел при
.
Оно определяется как предел при ![]() , стремящемся к бесконечности, выражения
, стремящемся к бесконечности, выражения
![]() . При
. При ![]() это выражение
очень близко к
это выражение
очень близко к ![]() , поэтому через 50 лет отношение численности наших бабочек
(Б/А) будет
, поэтому через 50 лет отношение численности наших бабочек
(Б/А) будет
Мы видим, что даже столь ничтожное, казалось бы, отличие (здесь 1/50) может
за обозримое время 50 поколений привести к заметным последствиям, изменению
отношения численностей почти в три раза, а через сто лет - в
![]() раза и далее в геометрической прогрессии: каждые пятьдесят лет - в
раза и далее в геометрической прогрессии: каждые пятьдесят лет - в ![]() раз.
раз.
Полезный способ оценки: если относительное преимущество в коэффициенте
размножения около ![]() , то через n поколений отношение численностей изменится примерно в
, то через n поколений отношение численностей изменится примерно в
![]() раз. Подчеркнем, что это просто оценка, в реальности сплетается много
разных причин успешного выживания и воспроизводства.
раз. Подчеркнем, что это просто оценка, в реальности сплетается много
разных причин успешного выживания и воспроизводства.
Повторим для березовой пяденицы тот же вопрос, что и для крабов: откуда взялась темная форма? Почему она вытесняет светлую, понятно, но как же объяснить ее появление? А если она в малых количествах была и раньше, то как же она выжила на светлых стволах?
Приведенные в начале главы примеры приспособления к изменившимся условиям допускают в первом приближении такое описание. Особи (крабы, бабочки) отличаются по какому-то одному признаку (ширине головогруди, окраске). Эти различия передаются по наследству. Успех в выживании и оставлении потомства зависит от них. Выразив эту зависимость количественно, можно предсказать, как будет меняться соотношение между численностями носителей различных вариантов признака.
Такая модель - резкое упрощение изложенной в предыдущей главе.
Рассматривается не биосфера, а одна популяция или, в крайнем случае,
небольшое число взаимодействующих популяций. Пространство ![]() - совокупность
возможных вариантов (значений) признака. Обычно
- совокупность
возможных вариантов (значений) признака. Обычно ![]() или конечно, или (для
непрерывных признаков) представляет собой отрезок прямой. Отличия по другим
признакам предполагаются несущественными. Для полового размножения надо еще
указать вероятности скрещивания носителей разных вариантов признака и
расщепление признака и потомства (описать генетику признака). В первом
приближении, однако, эффекты полового размножения часто не учитываются. Мы
тоже так поступили при оценке роли покровительственной окраски.
или конечно, или (для
непрерывных признаков) представляет собой отрезок прямой. Отличия по другим
признакам предполагаются несущественными. Для полового размножения надо еще
указать вероятности скрещивания носителей разных вариантов признака и
расщепление признака и потомства (описать генетику признака). В первом
приближении, однако, эффекты полового размножения часто не учитываются. Мы
тоже так поступили при оценке роли покровительственной окраски.
По отношению к реальности такие модели стоят заведомо не выше пятого типа (эвристическая модель без количественного подтверждения). Их достоинство - простота анализа. Иногда они все же хорошо согласуются с реальностью и, тем самым, переходят в четвертый тип (``опустим для ясности некоторые детали'').
Не станем приписывать существ, эволюцию которых мы будем изучать, к какому-нибудь биологическому виду. Отнесем их к царству матемазавров, хотя некоторые интересные черты реальности их эволюция все же моделирует.
Первая модель предназначена для понимания преднамеренных случайностей в фенотипе как приспособления к случайной среде. Она откровенно относится к восьмому типу - демонстрация возможности.
Рассмотрим организм, который может находиться только в двух существенно
различающихся состояниях - защищенном и незащищенном. Пусть состояния среды
тоже могут быть только двух разных типов - агрессивного и нормального.
Предположим также, что по прошествии некоторого фиксированного отрезка
времени (примем его за единицу измерения) состояние среды случайно меняется.
Пусть (упрощать так упрощать) новое состояние никак не зависит от
предыдущего, вероятность появления нормального состояния - ![]() , а агрессивного
-
, а агрессивного
- ![]() .
.
Замечание для читателя, не знающего, что такое вероятность:
сказанное означает, что из большой последовательности случаев
обычно в ![]() -й части появится нормальное состояние, а в
-й части появится нормальное состояние, а в ![]() -й -
агрессивное; если
-й -
агрессивное; если ![]() , то это можно понять так: каждый
раз бросается монетка, орел - нормальное состояние, решка -
агрессивное. Всегда
, то это можно понять так: каждый
раз бросается монетка, орел - нормальное состояние, решка -
агрессивное. Всегда
![]() есть много
элементарных руководств по теории вероятностей. Из них рекомендуем
книжку [33].
есть много
элементарных руководств по теории вероятностей. Из них рекомендуем
книжку [33].
Опишем взаимоотношения организма со средой. Если незащищенный организм
попадает в агрессивную среду, то он погибает. Если защищенный организм
попадает в агрессивную среду, то он переживает это без всякого ущерба, но не
размножается. Если незащищенный организм попадает в нормальную среду, то он
размножается и за единицу времени вместо одного такого организма получаем
![]() . Если защищенный организм попадает в нормальную среду, то он переходит в
незащищенное состояние и размножается, однако смена состояния требует
ресурсов и времени, поэтому в результате размножения из одного организма за
выделенный период времени получаем
. Если защищенный организм попадает в нормальную среду, то он переходит в
незащищенное состояние и размножается, однако смена состояния требует
ресурсов и времени, поэтому в результате размножения из одного организма за
выделенный период времени получаем ![]() все организмы рождаются
незащищенными. Обратным влиянием организмов на среду пренебрегаем. Описанные
взаимоотношения организма и среды могут быть наглядно представлены таблицей.
все организмы рождаются
незащищенными. Обратным влиянием организмов на среду пренебрегаем. Описанные
взаимоотношения организма и среды могут быть наглядно представлены таблицей.
=
table
tabular
|p142pt|p142pt|p142pt|
Пусть организмы отличаются друг от друга по одному наследуемому признаку. Он
состоит в наличии ``датчика случайных чисел'': по окончании единичного
времени, в течение которого среда была нормальной, перед случайной сменой
состояния организм с вероятностью ![]() переходит в защищенное состояние, а с
вероятностью
переходит в защищенное состояние, а с
вероятностью ![]() остается в незащищенном. Число
остается в незащищенном. Число ![]() - наследуемый
признак. Изучается отбор по этому признаку.
- наследуемый
признак. Изучается отбор по этому признаку.
Нужно еще добавить, что в период нормального состояния среды действуют
факторы смертности, не зависящие от состояния организма. Они меняют
соотношения численностей организмов, отличающихся по указанному признаку, но
поддерживают полную численность в некоторых пределах. Поскольку нас будет
интересовать именно соотношение численностей носителей различных значений
![]() , эти факторы явно в рассмотрение вводить не будем.
, эти факторы явно в рассмотрение вводить не будем.
Пусть нам уже задана свершившаяся история - последовательность состояний
среды за довольно большое время. Пусть в этой последовательности ``все, как
положено'' - ![]() я часть состояний нормальна,
я часть состояний нормальна, ![]() я агрессивна; за нормальным
состоянием в
я агрессивна; за нормальным
состоянием в ![]() й части случаев следует нормальное, а в
й части случаев следует нормальное, а в ![]() й - агрессивное. Такие
последовательности наиболее вероятны. Здесь выражение ``в
й - агрессивное. Такие
последовательности наиболее вероятны. Здесь выражение ``в ![]() й части'' надо
понимать с добавлением ``достаточно точно''. Отметим, что в случайных
последовательностях могут появляться очень длинные конечные отрезки, где все
далеко не ``как положено'', например, 100 решек подряд при бросании монеты.
Однако, чем длиннее последовательности решек, тем меньше вероятность такого
уклонения. Пусть для определенности первое состояние - нормальное. Примем
еще, что особей очень много, так что вместо вероятностей можно говорить о
частотах, долях.
й части'' надо
понимать с добавлением ``достаточно точно''. Отметим, что в случайных
последовательностях могут появляться очень длинные конечные отрезки, где все
далеко не ``как положено'', например, 100 решек подряд при бросании монеты.
Однако, чем длиннее последовательности решек, тем меньше вероятность такого
уклонения. Пусть для определенности первое состояние - нормальное. Примем
еще, что особей очень много, так что вместо вероятностей можно говорить о
частотах, долях.
Будем сравнивать между собой несколько различных значений ![]() .
Пусть к концу первого периода имелось
.
Пусть к концу первого периода имелось ![]() особей, для которых
особей, для которых
![]() - вероятность перейти в защищенное состояние (значение
признака). Сколько их будет к концу ближайшего периода с
нормальным состоянием среды? Если этот ближайший период -
следующий, то получим такую картину:
- вероятность перейти в защищенное состояние (значение
признака). Сколько их будет к концу ближайшего периода с
нормальным состоянием среды? Если этот ближайший период -
следующий, то получим такую картину: ![]() особей перейдет в
защищенное состояние,
особей перейдет в
защищенное состояние, ![]() останется в незащищенном:
первые дадут к концу следующего периода
останется в незащищенном:
первые дадут к концу следующего периода ![]() потомков, вторые
-
потомков, вторые
- ![]() , а всего
, а всего
![]() . Если же между
первым периодом и ближайшим к нему с нормальным состоянием среды
будут периоды с агрессивным состоянием (неважно, один или
несколько), то число потомков к концу этого ближайшего периода
будет другим:
. Если же между
первым периодом и ближайшим к нему с нормальным состоянием среды
будут периоды с агрессивным состоянием (неважно, один или
несколько), то число потомков к концу этого ближайшего периода
будет другим: ![]() особей перейдут в защищенное состояние,
остальные погибнут, и в итоге получим
особей перейдут в защищенное состояние,
остальные погибнут, и в итоге получим ![]() потомков.
потомков.
Пусть в рассматриваемой последовательности есть ![]() нормальное состояние
среды. Выясним, сколько особей со значением признака
нормальное состояние
среды. Выясним, сколько особей со значением признака ![]() будет к концу
последнего такого состояния. Для этого нужно
будет к концу
последнего такого состояния. Для этого нужно ![]() раз умножить
раз умножить ![]() на
на
![]() и
и ![]() раз - на
раз - на ![]() . Итак, получаем, что к концу
последнего нормального состояния число тех особей, для которых вероятность
перехода в защищенное состояние
. Итак, получаем, что к концу
последнего нормального состояния число тех особей, для которых вероятность
перехода в защищенное состояние ![]() , будет равно
, будет равно
таким образом оказывается, что число носителей значения признака ![]() изменяется
примерно в геометрической прогрессии со знаменателем
изменяется
примерно в геометрической прогрессии со знаменателем
![]()
Учет факторов смертности, не зависящих от ![]() , приведет к тому, что в
геометрической прогрессии будет меняться не число носителей значения
, приведет к тому, что в
геометрической прогрессии будет меняться не число носителей значения ![]() , а их
отношение для двух разных
, а их
отношение для двух разных ![]() . Все равно, со временем в указанных
предположениях сохраняются только носители того значения
. Все равно, со временем в указанных
предположениях сохраняются только носители того значения ![]() , для которых
знаменатель максимален. Найдем это значение. Нуль производной знаменателя по
, для которых
знаменатель максимален. Найдем это значение. Нуль производной знаменателя по
![]() отыскивается мгновенно:
отыскивается мгновенно:
![]() (упражнение!). но
(упражнение!). но ![]() по смыслу
не может быть больше 1. Поэтому после дополнительной проверки получаем ответ
для точки максимума знаменателя:
по смыслу
не может быть больше 1. Поэтому после дополнительной проверки получаем ответ
для точки максимума знаменателя:
если
![]() то
то
![]() ;
;
если
![]() то
то ![]()
Итак, в случайной среде особи со встроенным ``датчиком случайных чисел'' могут получать преимущества в темпе успешного размножения и, тем самым, вытеснять всех остальных.
С другой стороны, не всегда случайность среды автоматически приводит к
полезности случайного изменения фенотипа. В приведенной простейшей модели,
если ![]() то выгодно всем особям переходить в защищенную форму.
Напомним, что
то выгодно всем особям переходить в защищенную форму.
Напомним, что ![]() - вероятность появления агрессивного состояния среды,
- вероятность появления агрессивного состояния среды, ![]() - число потомков незащищенной особи в нормальной среде,
- число потомков незащищенной особи в нормальной среде, ![]() - число потомков
защищенной особи в нормальной среде. Оптимальное значение
- число потомков
защищенной особи в нормальной среде. Оптимальное значение ![]() растет при
увеличении
растет при
увеличении ![]() (вероятности агрессивного состояния среды),
(вероятности агрессивного состояния среды), ![]() (числа
потомков, даваемого защищенной особью после попадания в нормальную среду) и
убывает с ростом
(числа
потомков, даваемого защищенной особью после попадания в нормальную среду) и
убывает с ростом ![]() (числа потомков незащищенной особи в нормальной среде),
(числа потомков незащищенной особи в нормальной среде),
![]() Если бы выполнялось обратное неравенство
Если бы выполнялось обратное неравенство ![]() , тогда было бы
выгодно всем и всегда переходить в защищенное состояние. (Снова ``выгода''!
Трудно избежать этого слова, но здесь смысл кажется ясным.)
, тогда было бы
выгодно всем и всегда переходить в защищенное состояние. (Снова ``выгода''!
Трудно избежать этого слова, но здесь смысл кажется ясным.)
Итак, модель восьмого типа. На ней, однако, можно продемонстрировать ходы, характерные для рассматриваемого класса моделей ``отбор по признаку'':
Каждому значению наследуемого признака сопоставляется геометрическая прогрессия роста. Знаменатель этой прогрессии - ``мальтузианский параметр'' зависит от среды и значения признака. Он считается мерой оптимальности по принципу: у кого он больше, тот всех вытеснит.
Среда рассматривается как нечто внешнее и заданное, не зависящее от организмов. Никакой экологии.
Никакой генетики. Носители различных значений признака рассматриваются так, как будто они размножаются раздельно, не скрещиваясь. (Это важное следствие первого упрощения.)
Частичным основанием для 1, 2 может служить представление о разделении
факторов. Рассматривается отношение численности организмов, несущих значение
признака ![]() через шаг времени, к исходной их численности. Предполагается,
что это отношение с достаточной точностью можно представить произведением:
через шаг времени, к исходной их численности. Предполагается,
что это отношение с достаточной точностью можно представить произведением:
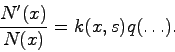
Здесь ![]() - начальное значение численности,
- начальное значение численности, ![]() - численность
носителей
- численность
носителей ![]() через шаг времени,
через шаг времени, ![]() - состояние среды,
- состояние среды, ![]() зависит только
от значения признака
зависит только
от значения признака ![]() и от состояния среды,
и от состояния среды, ![]() зависит от чего угодно (в
частности - от всех численностей) с одним ограничением: во все отношения
зависит от чего угодно (в
частности - от всех численностей) с одним ограничением: во все отношения
![]() (при всех значениях
(при всех значениях ![]() при данных состоянии среды и
распределении численностей должно войти только одно число
при данных состоянии среды и
распределении численностей должно войти только одно число ![]() не зависит
от
не зависит
от ![]() .
.
Числа ![]() участвуют в модели, числа
участвуют в модели, числа ![]() - в обосновании (для очистки
совести). С их помощью можно вроде бы учесть много факторов, зависящих от
состояния среды и всех численностей
- в обосновании (для очистки
совести). С их помощью можно вроде бы учесть много факторов, зависящих от
состояния среды и всех численностей ![]() , но не влияющих на отбор. Не
влияющих, так как они для всех
, но не влияющих на отбор. Не
влияющих, так как они для всех ![]() одинаковы и при сравнении численностей
для разных значений признака
одинаковы и при сравнении численностей
для разных значений признака ![]() и
и ![]() получим
получим
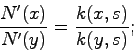
число ![]() сократилось. Поэтому доля носителей признака будет определяться
исключительно числами
сократилось. Поэтому доля носителей признака будет определяться
исключительно числами ![]() .
.
Экология может частично учитываться с помощью ![]() . Таким образом, экологии
не то, чтобы нет. Предполагается, что отбор и экология ``разделились''.
Подчеркнем, что под экологией мы подразумеваем не просто зависимость
организма от среды - такая зависимость явно входит в мальтузианский параметр
. Таким образом, экологии
не то, чтобы нет. Предполагается, что отбор и экология ``разделились''.
Подчеркнем, что под экологией мы подразумеваем не просто зависимость
организма от среды - такая зависимость явно входит в мальтузианский параметр
![]() , а их взаимную зависимость.
, а их взаимную зависимость.
Подробный разговор о генетике ждет нас впереди.
Теперь - одно формальное упрощение, которое было спрятано в наших рассуждениях. Оно не относится по своему существу к эволюционной тематике, но сыграло и играет большую роль в статистической физике. Поэтому скажем несколько фраз о нем.
Рассматривая существование в случайной среде, мы выбрали последовательность
состояний с наиболее вероятными характеристиками. Исходными были те же
соображения, что и при бросании монеты: наиболее вероятно, что орлов и решек
было поровну - в среднем-то их поровну. Итак: переход от среднего к наиболее
вероятному. Разбиралось существование организмов и отбор для
последовательностей с выбранными характеристиками. Отсюда был получен вывод:
в среднем наиболее успешно размножаются и выживают носители такого-то
значения признака. Рассуждения ``в среднем'' требуют, однако, совсем
другого. Для каждого ![]() нужно рассмотреть все возможные последовательности
агрессивных и нормальных состояний среды данной длины и найти, сколько в
среднем особей придется на одну начальную после такой последовательности.
Результаты потом надо усреднить по всем последовательностям: взять найденные
величины, умножить на вероятности последовательностей и сложить. Результат
получится другой. В приведенном примере его нетрудно точно вычислить, однако
ответ будет более громоздким.
нужно рассмотреть все возможные последовательности
агрессивных и нормальных состояний среды данной длины и найти, сколько в
среднем особей придется на одну начальную после такой последовательности.
Результаты потом надо усреднить по всем последовательностям: взять найденные
величины, умножить на вероятности последовательностей и сложить. Результат
получится другой. В приведенном примере его нетрудно точно вычислить, однако
ответ будет более громоздким.
Итак, два приема упрощения: среднее значение заменяется наиболее вероятным и среднее значение функции заменяется функцией от среднего значения. Эти приемы могут давать заметные ошибки, но в качестве первого приближения бывают полезны. Совсем простым аналогом может служить известное несовпадение в общем случае средней скорости со средним арифметическим скоростей на отрезках пути.
Насколько часто генетически запрограммированные случайности встречаются в природе? Существуют различные мнения. Так, Ф. Н. Семевский и С. М. Семенов полагают [90], что распространенное явление - диапауза (задержка развития у части особей) - во многом связано с этим эффектом.
Пример диапаузы: у растений одновременно созревшие семена могут прорастать не все сразу, а со сдвигом на год и более; у некоторых насекомых, проходящих различные фазы развития, часть особей задерживается в отдельных фазах. Это может рассматриваться как приспособление к случайностям в изменении среды со временем. Каким будет следующий год заранее неизвестно, поэтому может быть выгодно (ведет к увеличению коэффициента размножения) по-разному распределить свое потомство, чтобы оно развивалось несинхронно.
Кроме гипотетической случайности запрограммированной, внутренней, существует много случайностей внешних. Условия различны не только в разное время, но и в разных местах, неравномерно расселение многих видов и многое другое. Все это тоже может менять сроки развития. Поэтому дискуссия об основных причинах диапаузы закончится не скоро.
Не раз и не два приходилось нам слышать и читать истории о скромных хищниках. Суть их сводится к тому, что хищник пытается ограничить свое потребление, предпочитает питаться больными особями, не трогает беременных самок и молодняк жертв, может закусывать собственными детьми и т.п. и все это для того, чтобы, не дай бог, не подорвать популяцию жертвы - ведь в этом случае целиком погибнет и популяция хищника. На первый взгляд правдоподобно: умеренность выгодна, а неумеренность пагубна. Но понятие ``выгода'' сильно связано с человеческим пониманием этого слова, с умением предвидеть дальнейшие последствия поступков. (Человек, кстати, по отношению к природе часто ведет себя далеко не как скромный хищник, а намного менее выгодным образом).
Проведем мысленный эксперимент. Представим себе территорию, где скромно охотятся скромные хищники. Вдруг появляется ``суперхищник'', который не ограничивает охоту, интенсивно размножается, в общем - увеличивает на первых порах свой коэффициент размножения. Скромный хищник отстает в размножении, доля суперхищника увеличивается, поэтому можно сказать, что стратегия суперхищничества выгодна. Однако со временем, вытеснив окончательно скромных хищников и основательно расплодившись, суперхищник может уничтожить всех жертв и вымереть сам. А это невыгодно.
Итак, одно и то же поведение может быть и выгодно и невыгодно: свободное использование представлений о выгоде приводит к парадоксам.
Существуют различия между проявившимися в мысленном эксперименте выгодами скромного хищничества и суперхищничества. Суперхищник легко внедряется и вытесняет скромного хищника. Последний может длительно процветать (если не будет внедряться суперхищник). Увы, выгоды суперхищничества сказываются быстрее его недостатков: сначала внедряется, вытесняет скромного, а лишь потом гибнет. Ведущая к процветанию выгода скромности не защищена от внедрения.
Три важных свойства, совершенно различных по динамическим последствиям, склеились в рассмотренной ситуации под одним именем ``выгода''.
Устойчивое существование в отсутствие внедрения каких-либо разновидностей. Этим свойством обладает скромный хищник.
Устойчивость к внедрению новых разновидностей. Ею скромный хищник не обладает.
Способность внедряться, присущая суперхищнику.
Не сказано, обладает ли суперхищник первыми двумя свойствами, а скромный хищник - третьим. Это не случайно - сможет ли суперхищник устойчиво существовать, зависит от других персонажей - жертв. Если сможет, тогда имеет смысл говорить для него об устойчивости к внедрению, а без этого нельзя: о каком внедрении к суперхищнику может идти речь, когда не обеспечено его длительное существование? Если он все же обладает первым свойством, тогда наверняка будет устойчив к внедрению скромного хищника, а тот, соответственно, будет неспособен внедряться.
Чтобы длительно существовать в данных условиях, нужно обладать первыми двумя свойствами, а чтобы в них возникнуть - еще и третьим.
Первое свойство часто называют внутренней устойчивостью, второе - внешней. Но что устойчиво? Странный вроде бы вопрос. Ведь сказано: хищники одной разновидности, живущие в одном месте, короче - популяция хищников. Мы представляли себе такую ситуацию: есть хищники и жертвы, численность которых зависит в первую очередь от охоты хищников, все остальное - условия, которые ни от чего не зависят. Это слишком грубое приближение, и целесообразно перейти к единицам другого уровня.
В предыдущей главе не раз говорилось, что динамика мира живого не может считаться автономной и даже хорошо обособляемой (с выделением воздействий на нее). Лучше выделяются из окружения биосфера и биосферные единицы - биогеоценозы. Используем результаты проведенного мысленного эксперимента для определения трех типов устойчивости биогеоценозов.
Внутрення устойчивость. Рассматривается идеализированная ситуация, когда нет никакого притока живых существ в рассматриваемый биогеоценоз извне и новые разновидности (``вариации'') не проявляются также и внутри биогеоценоза. ``Номенклатура'' живых существ предполагается тем самым фиксированной. Возмутим слегка состояние биогеоценоза, изменив численность входящих в него живых существ или воздействуя на косные элементы. В ответ на такое возмущение будет получен динамический отклик - состояние биогеоценоза будет как-то меняться и далее. Пусть оно все же будет оставаться вблизи начального (невозмущенного) при всех достаточно малых возмущениях. Скажем тогда, что биогеоценоз в этом состоянии обладает внутренней устойчивостью.
Внешняя устойчивость. Всякий биогеоценоз существует на некотором фоне иммиграции - притока живых существ извне - и внутренней изменчивости. Новые живые существа появляются в малых количествах и либо внедряются, размножаясь до заметной численности, либо не могут внедриться - тогда их численность определяется темпами иммиграции или изменчивости. Если никакая разновидность живых существ из фона не может внедриться и тем самым изменить состояние биогеоценоза, то скажем, что он обладает внешней устойчивостью.
Обсуждать внешнюю устойчивость биогеоценоза имеет смысл только тогда, когда он обладает внутренней. Иначе он просто не просуществует достаточно долго для осуществления экспериментов по внедрению и изменится сам по себе.
В определении внутренней устойчивости присутствует такая идеализация: биогеоценоз, очищенный от всех процессов изменчивости и иммиграции. Для определения внешней устойчивости добавлена еще одна идеализация: фон изменчивости и иммиграции, представленный своей номенклатурой. Каждому биогеоценозу сопоставляются эти два идеализированных объекта, а внешняя устойчивость определяется как особое соотношение между ними.
Предлагается процедура проверки внешней устойчивости, имеющая вполне операциональную форму: следует проводить эксперименты по внедрению организмов из фона в биогеоценоз, освобожденный от наследственной изменчивости и иммиграции. Фон представлен номенклатурой, которая может быть как дискретной (список наименований), так и непрерывной (шкалы непрерывных признаков). Положение в номенклатуре предполагается наследуемым - дети относятся примерно в ту же клеточку (точку), что и родители. Эксперимент по внедрению состоит в том, что берется группа особей, принадлежащих одной клеточке (точке) из номенклатуры фона и помещается в биогеоценоз (освобожденный от процессов изменчивости и иммиграции). Умножит она свою численность и заметно повлияет на биогеоценоз - нет устойчивости. Вымрет, не увеличивая заметно своей численности, - надо брать другую группу и т. д. Не размножится ни одна - есть устойчивость.
Число особей во внедряемых группах должно быть достаточным для размножения, но все же малым13 - так, чтобы на первых порах они мало меняли состояние биогеоценоза и обратным влиянием (рикошетом) этих изменений на выживание и размножение внедренной группы можно было бы поначалу пренебречь. Пусть это выполнено, тогда размножение внедренной группы будет сначала происходить в геометрической прогрессии. Это позволяет количественно сформулировать условие внешней устойчивости.
Если знаменатель прогрессии - среднее число выживших потомков от одного предка за поколение (коэффициент размножения, он же - мальтузианский параметр) - больше единицы, то происходит нарастание численности внедренной группы. Рост может остановиться только тогда, когда воздействие размножившейся группы на биогеоценоз станет велико и состояние его изменится настольк, что обратное влияние (рикошет) будет существенным. В этом случае внешней устойчивости нет. Если знаменатель прогрессии меньше единицы, то внедренная группа просто вымирает, не оказав заметного воздействия. Получив такой результат для всех возможных групп, взятых из фона (и не представленных в биогеоценозе), можно сказать, что внешняя устойчивость есть.
Любому внутренне устойчивому состоянию биогеоценоза соответствует функция на номенклатуре изменчивости, сопоставляющая каждой ее клеточке (точке) коэффициент размножения соответствующей малой группы, внедренной в биогеоценоз. Удобно включить в номенклатуру изменчивости клеточки (точки) для тех живых существ, которые представлены в биогеоценозе. Этим клеточкам (точкам) соответствует единичный коэффициент размножения - ведь соответствующие живые существа длительно присутствуют в биогеоценозе (с исключенной иммиграцией и изменчивостью), не вымирая и не устремляя свою численность к бесконечности. Как уже говорилось при изучении внешней устойчивости предполагается, что биогеоценоз внутренне устойчив.
Условия внешней устойчивости можно сформулировать в форме принципа оптимальности. Обозначим ![]() множество клеточек (точек) номенклатуры изменчивости, маленькими
множество клеточек (точек) номенклатуры изменчивости, маленькими ![]() будем
обозначать его точки,
будем
обозначать его точки, ![]() коэффициент размножения для данного
коэффициент размножения для данного ![]() . Состояние
биогеоценоза предполагается заданным.
. Состояние
биогеоценоза предполагается заданным.
Необходимое условие внешней устойчивости: для всех ![]() из
из ![]() выполнено
неравенство
выполнено
неравенство ![]() (именно меньше или равно: где-то меньше, а где-то
равно; в частности, равно для тех
(именно меньше или равно: где-то меньше, а где-то
равно; в частности, равно для тех ![]() , которые соответствуют живым существам,
представленным в биогеоценозе).
, которые соответствуют живым существам,
представленным в биогеоценозе).
Достаточное условие внешней устойчивости: для всех ![]() из
из ![]() выполнено
неравенство
выполнено
неравенство ![]() ; равенство
; равенство ![]() имеет место
имеет место ![]() для тех
для тех ![]() , которые
соответствуют живым существам, представленным в биогеоценозе.
, которые
соответствуют живым существам, представленным в биогеоценозе.
Типичная ситуация ``отбора по признаку'': те же геометрические прогрессии,
простой мальтузианский параметр - коэффициент размножения и другое, только
![]() может быть устроено сложнее. В первой главе мы говорили о необходимо
возникающей нелинейности, как о проблеме. Здесь удалось ее обойти за счет
специально сконструированной ситуации - эксперимента по внедрению малых
групп.
может быть устроено сложнее. В первой главе мы говорили о необходимо
возникающей нелинейности, как о проблеме. Здесь удалось ее обойти за счет
специально сконструированной ситуации - эксперимента по внедрению малых
групп.
Принцип оптимальности - принцип максимума коэффициента размножения. В устойчивом биогеоценозе представлены только те живые существа, для которых коэффициент размножения достигает максимального значения, а это значение - единица. Это - не простой принцип оптимальности. Само состояние биогеоценоза определяет коэффициенты размножения для внедряемых групп. От тех живых существ, что в нем присутствуют, зависит функция, точки максимума которой соответствуют этим живым существам. И еще: не простая оптимальность, а совместимая, стоит заменить какую-нибудь разновидность живых существ, и оптимальность других нарушится. Для описания этого эффекта придуман специальный термин - сооптимальность.
Образ великого экспериментатора - Природы, постоянно проводящего эксперименты по внедрению и отбрасывающего все неустойчивое, очень убедителен. Англоязычная шутка: GOD - Generator of Diversity (бог расшифровывается по буквам как генератор разнообразия) ([71], c. 58). Существуют даже попытки включить внешнюю устойчивость в определение биогеоценоза [90]. Это рискованно.
Возможна такая ситуация. Складывается биогеоценоз. Входящие в него
живые существа изменяют среду своего обитания и создают
благоприятные условия для внедрения других. Те в свою очередь
меняют среду и создают условия для внедрения третьих и т. Д. Этот
процесс может стремиться к неподвижному состоянию (оно называется
терминальным или климаксным), а может и
безостановочно продолжаться. Тогда точки максимума коэффициента
размножения ![]() безостановочно перемещаются по множеству
безостановочно перемещаются по множеству ![]() .
Темп перемещения пропорционален интенсивности иммиграции и
изменчивости. Обнаружив этот эффект на моделях в 1978 году, мы
назвали его эффектом дрейфа. Получены уравнения,
описывающие дрейф в предположении малой изменчивости (см.
[35]). Не существует принципиального запрета на такое
перемещение. Более того, с малой скоростью оно, скорее всего,
происходит весьма часто. А в этом случае нет внешней устойчивости
состояний. Возможен выход - исследовать устойчивость не состояний,
а траекторий. Переход для математической теории устойчивости
стандартный, но это ответвление может завести нас слишком далеко.
.
Темп перемещения пропорционален интенсивности иммиграции и
изменчивости. Обнаружив этот эффект на моделях в 1978 году, мы
назвали его эффектом дрейфа. Получены уравнения,
описывающие дрейф в предположении малой изменчивости (см.
[35]). Не существует принципиального запрета на такое
перемещение. Более того, с малой скоростью оно, скорее всего,
происходит весьма часто. А в этом случае нет внешней устойчивости
состояний. Возможен выход - исследовать устойчивость не состояний,
а траекторий. Переход для математической теории устойчивости
стандартный, но это ответвление может завести нас слишком далеко.
Строго говоря, рассмотрение траекторий, а не стационарных состояний необходимо всегда: сезонные колебания, изменения погоды и др. приводят к очевидной нестационарности. Выход известен - усреднение по времени. Тогда принцип максимума коэффициента размножения заменяется на принцип максимума среднего (по прошлому, например) коэффициента размножения.
Устойчивая реализуемость. Существуют два пути определения этой устойчивости. Оба опираются на одну идею: заменить живые существа в биоценозе на близкие из фона и посмотреть, вернется ли он к прежнему составу - произойдут ли обратные замены. Заместители берутся из фона примерно в тех же количествах, в каких существовали исходные существа. Кроме возвращения к исходному составу важно также, чтобы уклонения ``по дороге'' были не слишком велики. Фон изменчивости и иммиграции присутствует теперь не только в виде номенклатуры, но и непосредственно представляет материал для перестройки. Процессы иммиграции и изменчивость вновь включены в реальном времени.
Первый путь основывается сразу на представлении о ``близких формах''. На множестве клеточек и точек номенклатуры изменчивости и иммиграции вводится мера близости, наример, расстояние. Эксперимент по проверке устойчивости состоит в том, что всех особей биогеоценоза заменяют на близких, как бы одним махом переводя из одних клеточек (точек) в другие.
Второй путь исходит из представления о той же по сути замене, но как о результате ``естественного'' процесса. Пусть несколько изменились внешние условия и некоторые варианты организмов их фона внедрились в биогеоценоз, а часть ранее присутствовавших ушла в фон. Восстановим условия и последим за изменениями системы: восстановится прежнее состояние, будут ли сильны уклонения по дороге к нему?
Если в рассматриваемых случаях возврат к прежнему состоянию всегда происходит и уклонения по дороге не слишком велики, то имеет место устойчивая реализуемость.
Основанием для введения устойчивой реализуемости служит представление о том, что биогеоценозы не являются ``райскими садами'', сотворенными из ничего. Напротив, они получены из других, живые существа постоянно замещались. Существование и устойчивость процессов замещения, ведущих в данных условиях к рассматриваемому состоянию биогеоценоза - вот сущность понятия ``устойчивая реализуемость''.
Часто приходится слышать, читать и писать о том, что теория эволюции объясняет целесообразность живых существ. Да, она дает возможность говорить о целях и о выгодах там, где действуют естественные процессы. Важное основание для этого - возникающие принципы оптимальности, в первую очередь тот, который был сформулирован в условиях внешней устойчивости. Он опирается на рассмотрение эксперимента по внедрению. Чтобы попасть в летопись живых существ, нужно внедриться. Чтобы существовать достаточно долго, нужно быть защищенным от внедрения конкурентов.
Идея устойчивости к внедрению имеет близкие аналогии в физике: ``Слово ``неустойчивый'' применяется в термодинамике не совсем в таком смысле, как в механике. Предположим, что у нас есть пересыщенный раствор. Кристаллизация в нем может вообще не начаться, вещество может быть устойчивым в пределах некоторых изменений состояния. Однако раствор начинает быстро кристаллизоваться при введении в него крошечного кристалла, т. е. раствор может оказаться неустойчивым по отношению к внедрению в него кристаллической фазы. Точно так же перегретая вода может быть устойчивой к разнообразным процессам, но не по отношению к внедрению в нее пузырька пара'' (Дж. В. Гиббс ([30], c. 551).
Теория эволюции не только разрешает рассуждать о целях и выгодах. Она еще задает особую рамку для этих рассуждений. Важный элемент этой рамки - эксперимент по внедрению. Нельзя просто разговаривать о целях и выгодах, не обращаясь к динамическим моделям, не рассматривая мысленно ситуации внедрения и существования. Три типа устойчивости - три смысла пользы, и все три должны присутствовать с детальным указанием на то, какой смысл имеется ввиду в каждом конкретном рассуждении. Иначе начинается движение не в смыслах, а в словах - бессмысленное движение.
Говорят, например, о пользе летальных (смертельных) мутаций для вида. Да появись конкурент без таких мутаций, он бы вытеснил в два счета это регулирующееся леталями чудо-юдо.
А легенда о скромном хищнике? Что защищает его от внедрения суперхищника? Простейший ответ: ничего. Скромность, скорее всего, не стратегия, а отражение физических возможностей.
Хищники и жертвы эволюционируют вместе. Каждый заяц ``мечтал'' бы быть ``суперзайцем'', легко убежать от всех хищников, ничем не болеть. Если бы такой суперзаяц вдруг возник, то ничто бы не помешало его внедрению. Он бы уничтожил ту растительность, которую мог бы съесть и либо вымер бы, либо влачил бы жалкое существование вблизи небольших возобновляемых запасов растений. Каждый волк мечтал бы быть суперволком, увы, с подобными же последствиями после осуществления ``мечты''. Нет запретов, делающих возникновение суперволков и суперзайцев невозможным. Кто помогает хищнику стать ``супер''? Жертва. А жертве - хищник. Так они вместе и эволюционируют. Кто забежит слишком вперед, тот скорее всего погибнет и, возможно, утащит за собой другого.
Распространена точка зрения: организмы должны непрерывно совершенствоваться, эволюционировать: кто отстал, тот погиб. Ее иногда называют ``гипотезой Черной королевы'' (вспомните ``Алису в Зазеркалье'': там Черная королева объясняла Алисе, что оставаться на месте можно только тогда, когда быстро идешь). Однако и тот, кто ``забежал вперед'', тоже может погибнуть быстрее, уничтожив отставшую живую часть своей среды обитания.
Оставим эти метафорические разговоры про ``вперед - назад'' в эволюции и вернемся к скромному хищнику. Не был ли нами нарушен закон презумпции осмысленности, когда мы отбросили идею о скромном хищничестве? Она не соответствует нашей модели? Но мы и не пытались придумать модель для оправдания ``скромности''. Нарушена заявленная в первой главе цепочка понимания: обосновывать, объяснять, применять, а потом уже критиковать.
Поступим так. Зафиксируем описанный эксперимент по внедрению как первый и главный способ проверки на выгодность - невыгодность, целесообразность - нецелесообразность - решающий эксперимент. С другой стороны, не будем считать его выводы окончательными и попробуем представить себе, где полученные результаты могут быть неверны.
Попытаемся дополнить модель. Она проста, но получена не упрощением - мы не имели исходной подробной модели. Но теперь, накладывая модель на реальность, можно пытаться выделить главные моменты несовпадения, главные с точки зрения возможного объяснения скромности. Нам бросаются в глаза три таких момента. Рассматривался один биогеоценоз, одна популяция. В действительности вид существует как система связанных популяций - это первое. Эксперимент по внедрению рассматривался как искусственная ситуация: внедрили группу и смотрим, внедрили другую - и смотрим, а можно характеризовать процесс изменчивости и иммиграции его интенсивностью - это второе. Мы предполагали, что хищники относятся друг к другу безразлично и обращали главное внимание на отношение ``хищник - жертва''. А многие высшие хищники вступают между собой в территориальные взаимоотношения, образуют тесные группы (стаи, кланы...) со сложной иерархической структурой и т.п. (третье).
Используем первые два замечания для конструирования модели, в которой скромный хищник существует в сети слабо связанных популяций и это существование устойчиво по отношению к редкому внедрению суперхищников в отдельные популяции.
Пусть весь ареал (область) обитания вида разбит на далеко отстоящие друг от
друга небольшие местообитания, в которых живут популяции скромных хищников.
Предположим, что есть ряд характерных времен, отделенных друг от друга
отношением ``много больше''. Самое большое из них - время появления мутанта
- суперхищника. Обозначим его ![]() . Следующее - время переноса
генетического материала между популяциями
. Следующее - время переноса
генетического материала между популяциями ![]() . Время
. Время ![]() много
меньше, чем
много
меньше, чем ![]() , поэтому будем предполагать появление суперхищников в
двух популяциях на отрезке времени
, поэтому будем предполагать появление суперхищников в
двух популяциях на отрезке времени ![]() невероятным. Третье время
невероятным. Третье время ![]() требуется возникшему в отдельной популяции суперхищнику на то, чтобы
вытеснить скромного хищника, подорвать популяцию жертвы и самому вымереть.
Предполагается, что
требуется возникшему в отдельной популяции суперхищнику на то, чтобы
вытеснить скромного хищника, подорвать популяцию жертвы и самому вымереть.
Предполагается, что ![]() много меньше, чем
много меньше, чем ![]() . Это позволяет
пренебречь вероятностью переноса суперхищника в другую популяцию - каждый
раз он будет вынужден возникать заново. Еще одно предположение: популяция
жертвы восстанавливается после разрушений за время порядка
. Это позволяет
пренебречь вероятностью переноса суперхищника в другую популяцию - каждый
раз он будет вынужден возникать заново. Еще одно предположение: популяция
жертвы восстанавливается после разрушений за время порядка ![]() . Итак,
. Итак,
![]() (запись » означает ``много больше'').
(запись » означает ``много больше'').
В указанных предположениях скромный хищник может существовать очень долго. Временами в отдельных популяциях будет возникать суперхищник, уничтожать их, вымирать, потом эти популяции будут вновь заселяться скромными хищниками.
Внимание! Здесь снова присутствует эксперимент по внедрению и внешняя устойчивость, только в модифицированном виде. Учтена пространственная структура и приняты гипотезы о пространственно-временных особенностях процесса внедрения - он предполагается состоящим из редких и локализованных событий. ``Искусственное'' экспериментирование заменено ``естественным'' процессом. Для экспериментирования достаточно было номенклатуры фона, для естественного процесса появляются пространственно-временные характеристики.
Один из немногих внушающих доверие примеров скромного хищничества
приводит М. М. Камшилов ([51], c. 163-164). Некоторые морские
звезды прекращают питание на один-два месяца, дожидаясь, пока
разовьются личинки их жертв. Масса жертв за время перерыва
увеличивается в 500 раз и более. Небольшое количество
``нескромных'' хищников могло бы, питаясь личинками, очень быстро
(практически за один сезон) подорвать популяцию жертвы, заодно
погибнуть и утащить с собой популяцию скромных хищников. Время
![]() здесь очень мало. А это главное: неравенство
здесь очень мало. А это главное: неравенство
![]() весьма правдоподобно и без дополнительных предположений
(исключение - изолированные острова в океане, перенос
генетического материала между которыми для многих видов
практически не идет и потому изменчивость опережает этот процесс).
весьма правдоподобно и без дополнительных предположений
(исключение - изолированные острова в океане, перенос
генетического материала между которыми для многих видов
практически не идет и потому изменчивость опережает этот процесс).
Увы, модель со слабо связанными популяциями не решает в общем случае вопроса о возможности существования скромного хищника без учета такого взаимодействия животных, которое бы пресекало суперхищничество. Модель со слабо связанными популяциями играет на том, что возможны только крайние варианты: скромный хищник и суперхищник. Стоит допустить между ними непрерывный (или хотя бы очень плотный) ряд переходных стратегий, как сразу и в приведенной ситуации окажется, что со временем будет происходить сдвиг от скромного хищничества к суперхищничеству, все популяции будут поставлены на грань вымирания, а потом и вымрут. Поясним это. Пусть все хищники ведут себя ``скромно'' и есть некоторый запас возможностей слегка увеличить потребление. Тогда слегка ``нескромные'' хищники, увеличивая свое потребление, выиграют за счет остальных и со временем их потомство составит большинство популяции, хотя сама популяция, быть может, уменьшится. Поэтому для убедительности предпринятой попытки объяснения скромности надо, чтобы уже малый сдвиг в сторону нескромности приводил к быстрому вымиранию и, с другой стороны, давал бы очень мало выгод. Именно так обстоит дело в приведенном примере с морскими звездами.
Можно и дальше усложнять модели, делая их все более искусственными или, наоборот, приближаясь к реальности, но мы не будем этим заниматься. Кажется ясным, что за редким исключением скромный хищник не может существовать, если между хищниками нет достаточно сложной системы взаимоотношений, препятствующей внедрению суперхищника. Мы уделяем так много места проблеме ``скромности'' не потому, что она очень важна, это не так, а по причине исключительной важности проблем, на которые она нас выводит. Вот и сейчас, заговорив о системе взаимоотношений, поддерживающих скромность, мы подошли к проблеме альтруизма.
Когда возникает вопрос о механизмах взаимодействия, поддерживающих такое поведение особей, которое им выгодно с точки зрения каких-нибудь представлений о выгоде, это всегда в той или иной степени вопрос об альтруизме. В философской энциклопедии [107] дается такое определение: ``альтруизм (франц. altruisme, от лат. alter - другой) - этич. принцип, заключающийся в бескорыстной заботе о благе ближнего''; противопоставляется эгоизму. Термин ``А'' введен О. Контом.
Говорить об этических принципах у животных не представляется особенно осмысленным. Но можно ли не назвать самоотверженным поведение птицы, уводящей хищника от гнезда? Как оценить предупреждающие крики, которыми подвергшаяся нападению особь оповещает об опасности окружающих, но подвергает тем самым себя большему риску?
Такой ``альтруизм'' в мире животных вызывает удивление на фоне сложившегося представления об определяющей роли ``инстинкта самосохранения''. Если попытаться проанализировать историю этих представлений, то, скорее всего, окажется, что они возникли под влиянием чисто человеческих воззрений на самоценность отдельной личности. Но если верить теории отбора, то важна не особь, а широко понимаемая репродуктивная способность - способность давать в достаточных количествах жизнеспособное и реально выживающее потомство, причем не только в одном поколении, а в их длинной последовательности.
Договоримся называть альтруизмом в биологии то, что кажется таковым на основании чисто человеческих представлений. Такой альтруизм распадается на несколько различных по своим причинам явлений.
Кажущийся альтруизм - поведение, которое воспринимается в силу незнания особенностей данного вида, а при детальном рассмотрении оказывается примитивно выгодным для особи. ``Весьма привлекательным для ряда исследователей оказался тезис об исключительном ``альтруизме'' рабочих муравьев, самоотверженно и без всякого вознаграждения денно и нощно пекущихся о товарищах по гнезду и даже чужих яйцах, личинках, куколках...
``Альтруизм'' рабочих муравьев также не выдерживает критики. Все формы ухода муравьев друг за другом и за молодью базируются на обязательной взаимопомощи как вознаграждении. Кормя личинку, муравей получает от нее капельку насыщенной ферментами пищи'' ([49], c. 21-22).
Таким образом, альтруизм рабочего муравья оказывается эгоизмом особи. Но тут происходит интересный поворот: согласно теории отбора в особом объяснении нуждается не тот ``альтруизм'', благодаря которому гены, принадлежащие особи, успешно размножаются, а как раз эгоизм особи. Муха, отложившая яйца, резко уменьшает свое стремление к самосохранению, и это понятно.
Эгоизм рабочего муравья может быть осмыслен с точки зрения отбора как инструмент для реализации ``альтруистических'' действий, способствующих успешному размножению его генов. Тут же надо сказать, что этот инструмент иногда подводит - достаточно появиться в муравейнике жучкам-ламехузам, выделяющим особо привлекательные для муравьев вещества, как личинки и куколки забрасываются и все начинают ухаживать за ламехузами.
Второй вид альтруизма - генетический эгоизм. Помогая родственникам, особь способствует размножению и выживанию носителей собственной генетической информации. Классический пример - защита потомства. ``Одной из лучших иллюстраций альтруизма такого же типа служит поведение вирусоподобной самовоспроизводящейся частицы, называемой плазмидой, которая паразитирует в бактериальных клетках. Время от времени одна из плазмид вырабатывает токсин, убивающий бактерию-хозяина и вероятно, саму плазмиду. Когда клетка-хозяин гибнет, токсин высвобождается, но убивает только тех из находящихся поблизости бактерий, которые не содержат плазмид. Бактериальные клетки, содержащие плазмиды, не повреждаются, потому что каждая плазмида вырабатывает кроме того, иммунопротеин, предохраняющий ее от действия токсина других плазмид. Таким образом, убивая бактерий-конкурентов, ген-самоубийца, продуцирующий токсин, способствует выживанию тех бактерий, которые содержат его генетические копии. Справедливость подобного истолкования подтверждается тем, что плазмиды обычно вырабатывают токсин при перенаселенности и конкуренции в культуре бактерий'' ([119], c. 199-200). Разобранный ``эгоизм'' муравья служит его генетическому эгоизму.
Следующая разновидность альтруизма основана на взаимности: я жертвую чем-то, помогая тебе, твердо рассчитывая на такое же твое поведение по отношению ко мне и моим родственникам. Этот альтруизм так и называют взаимным, или, не переводя на русский язык, реципкорным.
``Три случая взаимного альтруизма, приведенные Трайверсом (Р. Л. Трайверс - автор концепции взаимного альтруизма - примеч. авт.), - это симбиоз, связанный с чисткой, предупреждающие крики птиц и взаимный альтруизм14 человека. Выработка альтруистического поведения на основе реципрокного альтруизма требует жесткой системы отбора против ``обманщиков'' - особей, которые извлекают пользу из действий других, но в ответ при сходных обстоятельствах таких же действий не совершают'' ([45], c. 340).
Дж. Мейнард Смит [119] приводит более убедительные примеры взаимопомощи у павианов, подтверждающие концепцию взаимного альтруизма. Павианы явно обращаются друг к другу за помощью, причем не к кому попало, а избирательно. Там же Дж. Мейнард Смит отмечает: ``Недостаток состоит в том, что в этой ситуации, по-видимому, нет возможности гарантировать себя от обмана. Что мешает самцу А принять помощь, а затем, когда настанет его черед, не оказать ее? Возможное объяснение состоит в том, что стереотипы поведения у павианов складывались в процессе эволюции определенным образом, так что животные оказывают помощь только тем особям, которые в свою очередь помогают им. В этом случае обман был бы невыгодным. Конечно, в основе этой гипотезы лежит предположение, что осби способны узнавать других особей и помнить об их поведении в прошлом, однако наличие у павианов подобных способностей вполне допустимо'' ([119], c. 205-206).
Конечно, ``альтруизм'', основанный на отношениях ``ты - мне, я - тебе'' и индивидуальном узнавании, не кажется невероятным у таких высокоразвитых животных, как павианы. Но вряд ли он широко распространен по всему миру живого. Представляется более вероятным, что организмы отличают ``своих'' (по отношению к которым проявляется альтруизм) от ``чужих'' по каким-то отдельным опознавательным признакам - меткам.
Замечательным примером метки является гнездовой запах (запах семьи) у муравьев. На основании этого запаха муравьи делят особей одного с ними вида на своих и чужих: по отношению к своим - взаимопомощь, к чужим - агрессивное поведение. Могут возникать даже затяжные войны, приводящие к гибели многих муравьев. Запах гнезда не является генетически предопределенной особенностью. Более того, индивидуальность может быть утрачена. Когда в семью северного лесного муравья, насчитывающую примерно 15000 особей, трижды с интервалом в день вводили по тысяче особей из других муравейников этого вида, агрессивность по отношению к новичкам каждый раз снижалась и третья их партия была принята относительно спокойно. ``Предыдущие массовые введения новых качеств настолько расшатали монолитность небольшой муравьиной семьи, что она оказалась не в состоянии отличать своих от чужих'' ([49], c. 76).
Образовывая отводок от материнского гнезда, муравьи тратят немало сил на сохранение родственных отношений. Родство меток - запахов поддерживается непрерывным обменом особями. Почкованием создаются колонии муравейников. Но не только почкованием! Чужие муравейники могут ``породниться'', наладив обмен особями. Это уже не взаимный альтруизм ``ты -мне, я- тебе'', основанный на индивидуальном узнавании, и не генетический эгоизм - своими друг другу становятся первоначально не родственные муравейники.
Но и это еще не все. Колонии могут объединяться в федерации. И снова путем обмена особями создается родство запахов. В книге А. А. Захарова [49] указывается, что федерация - не просто большая колония. Колонии сохраняют свою целостность и после объединения. Важна иерархическая структура: муравейники объединены в колонии, колонии - в федерации.
Встречаются не только объединения, но и распады. Муравейник может отделиться от своей колонии и довольно быстро потерять родство меток - запахов. Поддержание родства требует специальных затрат, а его разрушение при отсутствии обменов происходит автоматически.
Все эти процессы плохо укладываются в схему взаимного альтруизма, хотя отдаленно ее и напоминают. Наличие особых меток, динамичность системы взаимоотношений позволяет говорить еще об одном типе альтруизма - коалиционном.
Мало кто не знает простейшей карточной игры в ``дурачка''. Если играют трое ``каждый за себя'', то при одинаковом навыке доля проигрышей каждого игрока будет в среднем равна 1/3. Но если двое договариваются подыгрывать друг другу, то третий почти все время будет в дурачках. Такое взаимовыгодное объединение в теории игр называется коалицией.
Стандартная задача теории игр с коалициями, решение которой излагается во многих учебниках, состоит в следующем. Пусть в некоторой степени несколько промышленных компаний получили предложение заключить выгодный контракт на заданную сумму. Вопрос в том, как разделить эту сумму между участниками. Дополнительная сложность: некоторые участники, если раздел их не удовлетворит, могут объединиться в коалицию и заключить контракт на стороне. Пусть для каждой мыслимой коалиции указана сумма контракта, который она может заключить. Спрашивается, возможен ли такой раздел, чтобы образование коалиций не было бы выгодно, т.е. никакая группа не получила бы в результате заключения контракта на стороне большей суммы. Другой, более сложный и не всегда имеющий единственное решение вопрос: если такого раздела не существует и каждая участвующая в игре компания стремится максимизировать получаемую сумму, то каковы будут образовавшиеся коалиции?
В теории отбора: выигрыш - число потомков за много поколений, коалиция - сообщество с взаимно альтруистическим (в той или иной степени) поведением. Спрашивается, каковы будут коалиции и как они будут меняться со временем?
Вопрос далеко не простой, но допускающий четкую формализацию. В каждом конкретном случае нужна дополнительная информация о возможной природе меток, по которым члены коалиции могут отличать своих от чужих. Проще всеготот частный случай, который обсуждался при рассмотрении взаимного альтруизма: члены коалиции знают друг друга ``в лицо'' и признают кого-либо своим только при том условии, что он выполняет ``условия коалиции'' - ведет себя достаточно альтруистически. Но гнездовой запах муравьев дает пример другой метки, именно метки, условного знака.
В связи с использованием меток может возникнуть вопрос: почему в коалицию не внедряются ``эгоисты'', несущие ту же метку? Они же получают двойную выгоду: к ним будут относиться альтруистически, а сами они ничем жертвовать не будут.
Что же, резонно предположить, что при наличии изменчивости, рождающей хорошо замаскированных эгоистов достаточно часто, вид не способен к образованию устойчивых коалиций. Оставим в стороне вопрос о том, будет ли он вытеснен другим видом, подверженным такой изменчивости в меньшей степени. Этот вопрос заведет нас в дебри дискуссии о групповом отборе, для которого основными отбираемыми единицами служат надорганизменные системы. Приведем только простую модель восьмого типа - демонстрацию того, как возможно сосуществование эгоистов и альтруистов в нестационарном режиме.
Петля альтруизма - так мы назвали эту модель. Представим себе альтруистическую коалицию, занимающую некоторое местообитание. Все особи коалиции несут метку, позволяющую им отличить своих - членов коалиции - от чужих. По отношению к своим проявляется альтруизм, к чужим - эгоизм. Пусть такая стратегия обеспечивает наилучшее выживание потомства в последовательности поколений при условии одинакового поведения всех особей коалиции.
При длительном существовании начинается ``загнивание'' альтруизма, появляются хорошо замаскированные эгоисты - носители той же метки. Начиная с некоторого порога их численности альтруистическое поведение по отношению к носителям метки становится невыгодным - ведет к вымиранию. Альтруисты либо вымирают, либо меняют стратегию на эгоистическую. Через некоторое время может возникнуть небольшая локальная альтруистическая коалиция - носительница другой метки. Такое объединение дает его участникам преимущество в размножении и они вытесняют эгоистических особей (либо частично инкорпорируют - включают в свою коалицию). Такой процесс можно сравнивать с распространением пожара. Далее - снова ``загнивание''. И так далее...
сложно сказать, насколько часто петля альтруизма встречается в действительности. Но, без сомнения, динамика коалиций - один из важных факторов эволюции.
В изучении поведения всех живых существ традиционно используется немало терминов, первоначально возникших для описания сознательных действий человека: альтруизм, эгоизм... .''Как всегда, эти рассуждения не означают, что животные сознательно подсчитывают преимущества альтруистического поведения. Просто особи, ведущие себя определенным образом, оставляют в среднем больше потомков, и это приводит к распространению генов, обусловливающих такое поведение'' ([45], c. 340).
Построить теорию культуры на основании биологических законов невозможно, но поиск аналогий полезен, если не слишком увлекаться и не придавать им чрезмерно глубокого смысла. Особенно легко найти интересные аналогии для динамики коалиций и их меток - ведь само понятие ``коалиция'' пришло из науки, изучающей некоторые особенности человеческой деятельности - из математической экономики. Мы не можем удержаться от соблазна привести несколько классических цитат.
``Великие силы Поднебесного мира, долго будучи разобщенными, стремятся соединиться и после продолжительного соединения опять распадаются - так говорят в народе''. Этой фразой, которую легко истолковать как указание на динамику коалиций, начинается классический эпос Лу Гуань-чжуна ``Троецарствие'', созданный в XIV веке.
Если понимать основную мысль следующего отрывка из работы И. В. Киреевского как необходимость рождения этноса сразу вместе с ``меткой'', выделяющей ``своих'', то мы с этим согласны, но трудно согласиться с тем, что такая метка обязательно должна быть религиозно-мифологической.
``Мифы не могут быть изобретены, ибо тогда является вопрос: кто изобрел их? Кто-нибудь один не мог их выдумать, ибо мифология так срастается с народом, что почитать ее изобретенною то же, что выводить язык народа из чьего-либо сочинения. И как бы мог изобретатель передать свои понятия народу и заставить его верить, что его вымыслы суть настоящие боги? Легче даже предположить, что мифология есть изобретение целого народа, но такое поэтическое состояние трудно объяснить исторически. Впрочем, это предположение встречало себе противоречие в самом понятии о народе. Ибо что же такое народ и что делает собрание людей народом? Не совместное бытие многих отдельных людей составляет народ, но общность сознания, выражающаяся наружно в языке, а внутренне в общности одного взгляда на порядок вещей и мира, единомыслие, из которого происходят нравы и обычаи. Эта общность взгляда на порядок и первоустройство вещей есть не что иное, как религия, которая у древних народов является в образе мифологии. Поэтому несообразно думать, чтобы мифология примкнула после к народу, уже существующему, когда народ уже не может возникнуть без общего взгляда на первоустройство мира, без мифологии. Итак, если мифология не может возникнуть в народе уже существующем, то она необходимо должна возникнуть вместе с самим народом, и, следовательно, она есть не что иное, как тот участок всеобщего сознания, с которым народ вышел из первобытного единства человечества, чтобы существовать как какой-то определенный и от всех отдельный народ. Мифология народа ест то наследство, которое он получает из прежнего единства и с которым он отделяется от общности человечества'' ([54], c. 243-244).
Замечательный пример, напоминающий ``петлю альтруизма'', приводит Ф. Энгельс: ``Ислам - это религия, приспособленная для жителей Востока, в особенности для арабов, следовательно, с одной стороны, для горожан, занимающихся торговлей и ремеслами, а с другой - для кочевников-бедуинов. Но в этом лежит зародыш периодически повторяющихся столкновений. Горожане богатеют, предаются роскоши, проявляют небрежность в соблюдении ``закона''. Бедуины, которые живут в бедности и вследствие бедности придерживаются строгих нравов, смотрят на эти богатства и наслаждения с завистью и жадностью. И вот они объединяются под предводительством какого-нибудь пророка, махди, чтобы покарать изменников веры, восстановить уважение к обрядам и истинной вере и в качестве вознаграждения присвоить себе богатства вероотступников. Лет через сто они, естественно, оказываются точно в таком же положении, в каком были эти вероотступники; необходимо новое очищение веры, появляется новый махди, игра начинается сначала'' ([123], c. 470).
Можно приблизить ``петлю альтруизма'' к этой ситуации, предположив, что местообитание неоднородно и загнивание в одних частях происходит раньше, чем в других.
Настоящий альтруизм, наиболее соответствующий человеческому пониманию этого слова, альтруизм без разделения на своих и чужих в биологии неустойчив - носители такого поведения вымирают. По мнению А. С. Штерна одна из важных мыслей романа Ф. М. Достоевского ``Идиот'' - обреченность носителей истинного альтруизма из-за жестокости общественных отношений.
При объяснении альтруизма, не являющегося генетическим эгоизмом, приходится наряду с номенклатурой изменчивости (X) вводить еще набор меток, служащих для разделения на своих и чужих. Какую-то метку особь несет с рождения. В этом смысле метка наследуется, но не обязательно строго. Существуют возможности смены меток (по крайней мере, у некоторых видов). Процесс смены меток всегда имеет определенные правила, порождаемые природой метки и биологией вида. Стратегии коалиционного поведения можно изучать методом ``отбора по признаку'', исследуя, какой набор стратегий выживает в результате отбора. Сами модели динамики с учетом смены меток заведомо сложнее и намного менее изучены, чем простой отбор по признаку.
Наибольший недостаток всех моделей данной главы - почти полное пренебрежение генетикой полового размножения. Все рассуждения велись так, как будто организмы воспроизводятся целиком. Исходя из этого строилось множество X, и множество наследуемых значений признака. Модели отбора, в которых генотипы отдельных организмов представлены как воспроизводящиеся почти целиком, называются классическими в противовес балансовым моделям, в которых предполагается, что генотип каждый раз ``переконструируется'' из генов родителей. Неправильно будет считать, что классические и балансовые модели по-разному описывают один и тот же процесс, противореча друг другу. Они выделяют два разных процесса, а ``генотип'' классической модели нечто иное, чем ``генотип'' балансовой. Впереди еще будет повод обсудить соотношения классических и балансовых моделей.
С разнополым размножением связана одна важная и до сих пор
нерешенная проблема: почему оно существует? Известно, что возможен
переход к бесполому размножению. Есть виды, например, у ящериц,
состоящие только из самок, откладывающих яйца, из которых снова
выходят самки (партеногенез). Переход к такому
размножению дает со временем двукратное преимущество в темпе
размножения. Это быстро реализующееся преимущество подробно
разобрано Дж. Мэйнардом Смитом в духе моделей отбора по признаку:
``Предположим, что у вида, размножающегося половым путем и
насчитывающего равное число самцов и самок, возникает мутация, в
результате которой самка переходит к партеногенетическому
размножению и производит лишь себе подобных самок. Число яиц ![]() ,
откладываемых одной самкой, обычно не зависит от того,
партеногенетически она размножается или нет, а определяется лишь
тем, как много пищи она может усвоить сверх того количества,
которое необходимо ей для поддержания собственного существования.
Аналогичным образом вероятность
,
откладываемых одной самкой, обычно не зависит от того,
партеногенетически она размножается или нет, а определяется лишь
тем, как много пищи она может усвоить сверх того количества,
которое необходимо ей для поддержания собственного существования.
Аналогичным образом вероятность ![]() того, что возникшая из яйца
особь доживет до размножения, тоже обычно не зависит от характера
размножения'' ([76], c. 13). Пусть в начальный момент имелось
того, что возникшая из яйца
особь доживет до размножения, тоже обычно не зависит от характера
размножения'' ([76], c. 13). Пусть в начальный момент имелось
![]() взрослых партеногенетических особей и
взрослых партеногенетических особей и ![]() тех, что используют
раздельнополое размножение (из них
тех, что используют
раздельнополое размножение (из них ![]() самок). Отложение яиц у
партеногенетических самок kn, у размножающихся половым
способом - kN/2 до репродуктивного возраста доживет
партеногенетических особей Pkn, использующих половое
размножение - PkN/2. Отношение численностей
(партеногенетические / размножающиеся половым путем) возрастает
вдвое. В начальный момент оно равно n/N, а для следующего
поколения
самок). Отложение яиц у
партеногенетических самок kn, у размножающихся половым
способом - kN/2 до репродуктивного возраста доживет
партеногенетических особей Pkn, использующих половое
размножение - PkN/2. Отношение численностей
(партеногенетические / размножающиеся половым путем) возрастает
вдвое. В начальный момент оно равно n/N, а для следующего
поколения![]()
Таким образом, партеногенез дает колоссальную выгоду внедрения. Вопрос о том, что обеспечивает существование раздельнополого размножения (зачем нужны самцы?), является центральным в книге Дж. Мэйнарда Смита [76], однако он не считает эту проблему окончательно решенной.
Ясно одно: двукратное преимущество при внедрении партеногенеза не позволяет отделываться от проблемы существования раздельнополого размножения заявлениями об увеличении скорости эволюции и росте разнообразия. Нужны согласованные с опытными данными количественные оценки. В цитированной книге это ясно показано.
Задумавшись над отношением ``отбора по признаку'' к реальности мы неизбежно приходим к необходимости уточнения понятия ``признак''. Попробуем определить его как особенность фенотипа. В этом случае возникает проблема наследования: не все особенности фенотипа наследуются, например, вес тела человека в большой степени зависит от условий жизни. Для рассматриваемых же моделей отбора важны именно в той или иной степени наследуемые особенности.
Попытаемся улучшить определение, положив: признак - генетически предопределенная особенность фенотипа. Опять плохо - генетически предопределены чаще всего не особенности фенотипа, а зависимость фенотипа от условий развития. Проводя отбор по одному и ому же признаку фенотипа в разных условиях, селекционер, очевидно, получит разные результаты [51]. Если же учесть возможность случайной связи между этими условиями и фенотипом, то ситуация становится еще более запутанной: генетически предопределены лишь вероятности появления различных фенотипов в данных условиях.
Есть подход к определению признаков, идущий от легкости их описания исследователем. С этой точки зрения признак - простое (просто выражаемое одним числом и немногими словами) свойство фенотипа. Сюда относятся рост, вес, цвет, простые особенности характера и т.п. такая простота обычно подразумевается, когда речь идет об отдельном признаке. Но она же приводит и ко многим проблемам. То, что просто с точки зрения описания, не обязательно является элементарным, выделяемым в самостоятельную единицу, достаточно обособленную в ходе индивидуального развития и передачи наследственной информации. Речь идет о корреляции признаков, давно замеченной селекционерами. Ч. Дарвин на эту тему пишет: ``таким образом, человек, отбирая и накопляя какую-нибудь особенность строения, почти наверняка будет неумышленно изменять и другие части организма на основании таинственных законов корреляции'' [38]. ``Таинственных законов'' - не меньше.
Яркий пример корреляции получен сравнительно недавно при одомашнивании лисиц. Отбирали и скрещивали между собой лисиц, более дружелюбных по отношению к человеку, а они ``особачились''. И хвост завился колечком, и окраска стала пегой, и другие внешние признаки стали более собачьими. Воздействовали вроде бы на один признак - дружелюбие, а изменились многие. Такое всестороннее изменение рассматривается Д. К. Беляевым как пример ``дестабилизирующего отбора'' (подробнее см. в его статье [11]): ``Отбор становится дестабилизирующим тогда, когда под его влияние попадают... системы нейроэндокринной регуляции онтогенеза'' - индивидуального развития.
Хотя мы и не дали окончательного определения признака, обсуждение этого понятия с разных сторон делает его более ясным, а также выявляет проблемы, связанные с его использованием. Для модели ``отбор по признаку'' лучше всего подходят признаки, простые с точки зрения описания и хорошо используемые - генетически предопределенные в данном диапазоне условий.
Попытаемся понять место обсуждаемых моделей, используя аналогию с физикой. Простейшая задача механики - изучение движения частицы по прямой в потенциальном поле. Движение сложной системы может быть разложено на простые одномерные.
Движение по прямой уподобим отбору по признаку, а сложную систему - реальности. Настоятельно повторяемый в сложной ситуации вопрос: ``по какому признаку идет отбор?'' аналогичен требованию указать прямую, по которой движутся частицы в сложной механической системе. Такой признак (как и прямую) можно точно указать только в специальных случаях, а чаще всего - в искусственно созданных ситуациях.
Модели ``отбор по признаку'' хороши своей простотой и понятностью, но слишком упрощают суть дела и по отношению к реальной эволюции стоят где-то на уровне шестого (``учтем только некоторые особенности'') или пятого (эвристического) типов, если, конечно, не прибегать к необычному расширению содержания понятия ``признак''. Изредка они могут подниматься до четвертого типа (``опустим для ясности некоторые детали''), но вряд ли выше.
Это отдельные модели, а система таких моделей является основным смыслообразующим звеном теории эволюции. На первых шагах эволюционных рассуждений обычно так или иначе строятся классические модели отбора по признаку (и лучше, если это делается явно). Далее они накладываются на реальность, обнаруживаются несовпадения и моменты нестыкованности с имеющимся знанием. Критикуя, перестраивая и дополняя модель, можно выработать более полное понимание и сформировать научное знание об исследуемом процессе.